
安徽羽叶报春同型花和二型花居群的遗传多样性和遗传结构分析
2018-06-06 02:49李永权徐延年吕国胜邵剑文
植物资源与环境学报 2018年2期
李永权,章 伟,徐延年,吕国胜,邵剑文,,①
(1. 安徽师范大学生命科学学院,安徽 芜湖 241000;2. 安徽省重要生物资源保护与利用研究重点实验室,安徽 芜湖 241000)
二型花柱(distyly)是一种最常见的雌雄蕊异位类型,其产生和维持机制一直是进化生物学和生态学的研究热点,迄今为止,至少发现有23科植物存在二型花柱[1]。二型花不但具有明显的雌雄蕊交互式异位特征(即长柱花柱头位于花筒口且花粉囊位于花筒中部,短柱花柱头位于花筒中部且花粉囊位于花筒口),而且还具有严格的自交不亲和特性,导致二型花植物只能通过不同花间的受粉来完成正常结籽过程[2-4]。但是,偶尔发生的等位基因重组互换等现象可导致植物的二型花同型化,形成柱头和花粉囊等高的长柱同型花或短柱同型花,自交不亲和性随之丧失,这些二型花植物在开花后可自花受粉并结籽[5]。通常情况下,在二型花转变为同型花后,植物的繁育系统也由异交变为自交,这一转变可能会降低植物有效种群规模,增加连锁不平衡和基因清除效应,进而降低植物居群的遗传多样性,促进居群间的遗传分化[6],甚至可能产生新物种[7-8]。
全世界的报春花属(PrimulaLinn.)植物约有500种[9-10],其中,近90%的种类具有二型花柱,剩余约10%的种类具有同型花柱。报春花属植物是最早开展二型花柱适应性进化研究的植物材料[3],目前该属已成为二型花柱进化生物学研究的模式材料。安徽羽叶报春(PrimulamerrillianaSchltr.)具有典型的二型花柱,花期较长,园艺开发和应用价值很高[11-12]。然而,该种仅分布于安徽南部海拔50~1 000 m的丘陵山区,喜生于山体北坡比较阴湿的林下、溪边和路旁的岩壁上[13]。近几十年来,由于人类活动对生境的破坏作用,安徽羽叶报春的野生种群数量明显下降,因此,陈明林等[14]建议将其列为安徽省重点保护野生植物。
相关研究结果显示:安徽羽叶报春二型花居群的遗传多样性较高,且在不同生境中分化明显[15-16]。作者在2014年至2015年的野外调查中发现,安徽省铜陵市凤凰山和池州市阴山存在安徽羽叶报春的长柱同型花居群,并且,与相近区域的二型花居群个体相比,除花朵同型、花冠直径较小外,这些长柱同型花居群个体的其他形态性状几乎没有区别。然而,这些同型花居群与二型花居群的遗传分化和遗传多样性差异及基因流情况等均未明确。为了弄清这些问题,作者选用扩增结果稳定且扩增条带多态性较高的10对SSR引物,对分布于安徽省秋浦河流域铜陵市凤凰山和池州市阴山的安徽羽叶报春2个长柱同型花居群及其附近区域的4个二型花居群的总DNA进行了PCR扩增,在此基础上对6个居群的遗传多样性和遗传结构进行了分析,以期明确安徽羽叶报春二型花转变为同型花后遗传多样性和遗传结构的变化规律,为探究植物二型花柱的维持机制提供参考资料,并为安徽羽叶报春保护策略制订提供指导依据。
1 材料和方法
1.1 材料
于2016年3月至4月,对安徽省秋浦河流域的6个安徽羽叶报春居群进行采样,各居群的基本情况见表1。其中,铜陵市义安区凤凰山(FHS)和池州市贵池区阴山(YS)居群为同型花居群(均为长柱同型花,柱头与花粉囊等高且均位于花筒口),分别位于秋浦河流域下游的低矮丘陵山区;池州市石台县大王洞(DWD)、池州市贵池区谷村(GC)、池州市石台县立新村(LXC)和池州市石台县莲花田(LHT)居群为二型花居群(长柱花和短柱花比例约1∶1),其中,DWD和GC居群位于秋浦河流域中游的低山或丘陵区,LXC和LHT居群位于秋浦河流域上游的低山或丘陵区。每个居群随机选取约20株相对分散的植株,每株采集2或3枚长势相似的嫩叶,放入透气的茶叶包内,用硅胶密封干燥,于室温条件下保存、备用。
表1安徽羽叶报春供试居群的基本情况
Table1BasicstatusofpopulationsofPrimulamerrillianaSchltr.tested
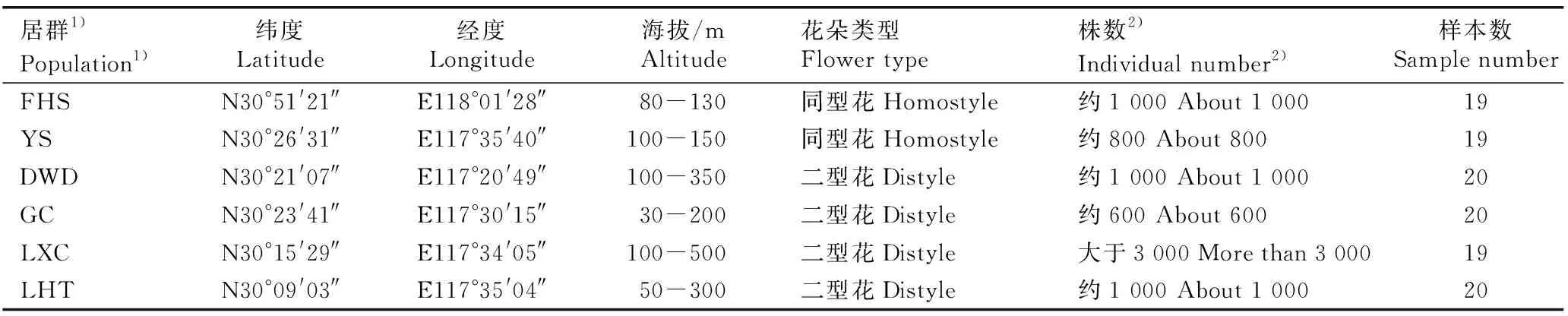
居群1)Population1)纬度Latitude经度Longitude海拔/mAltitude花朵类型Flower type株数2)Individual number2)样本数Sample numberFHSN30°51'21″E118°01'28″80-130同型花Homostyle约1 000 About 1 00019YSN30°26'31″E117°35'40″100-150同型花Homostyle约800 About 80019DWDN30°21'07″E117°20'49″100-350二型花Distyle约1 000 About 1 00020GCN30°23'41″E117°30'15″30-200二型花Distyle约600 About 60020LXCN30°15'29″E117°34'05″100-500二型花Distyle大于3 000 More than 3 00019LHTN30°09'03″E117°35'04″50-300二型花Distyle约1 000 About 1 00020
1)FHS:铜陵市义安区凤凰山 Fenghuangshan in Yian District of Tongling City;YS:池州市贵池区阴山 Yinshan in Guichi District of Chizhou City;DWD:池州市石台县大王洞 Dawangdong in Shitai County of Chizhou City;GC:池州市贵池区谷村 Gucun in Guichi District of Chizhou City;LXC:池州市石台县立新村 Lixincun in Shitai County of Chizhou City;LHT:池州市石台县莲花田 Lianhuatian in Shitai County of Chizhou City.
2)居群内安徽羽叶报春株数 Individual number ofPrimulamerrillianaSchltr. in population.
1.2 方法
采用改良的CTAB法[17]提取各居群嫩叶的总DNA。选用本实验室开发的扩增结果稳定且扩增条带多态性较高的10对SSR引物,各引物的序列及退火温度见表2。引物合成交由生工生物工程(上海)股份有限公司,并对上游引物的5′端进行荧光修饰(FAM、HEX或TAMRA)[15]。
扩增反应体系总体积为15.0 μL,包括DreamTaq Green PCR Master Mix(2×)7.5 μL、100 ng·μL-1总DNA 0.4 μL、100 μmol·L-1正向引物和反向引物各0.3 μL、ddH2O 6.5 μL。使用Bio-Rad iCycler PCR仪(美国Bio-Rad公司)进行扩增反应,扩增程序为:94 ℃预变性5 min;94 ℃变性30 s、引物相应退火温度退火60 s、72 ℃延伸90 s,35个循环;72 ℃延伸10 min。
扩增产物在质量体积分数2.0%琼脂糖凝胶上进行电泳检测;按照不同荧光混合后交由上海点晶生物科技有限公司采用ABI 3700基因测序仪(美国ABI公司)进行毛细管电泳测序分析,使用GeneMarker 1.91软件对测序结果进行判读,获得微卫星基因型数据。
表2用于安徽羽叶报春6个居群PCR扩增的引物序列及退火温度
Table2SequencesandannealingtemperaturesofprimersusedforPCRamplificationofsixpopulationsofPrimulamerrillianaSchltr.
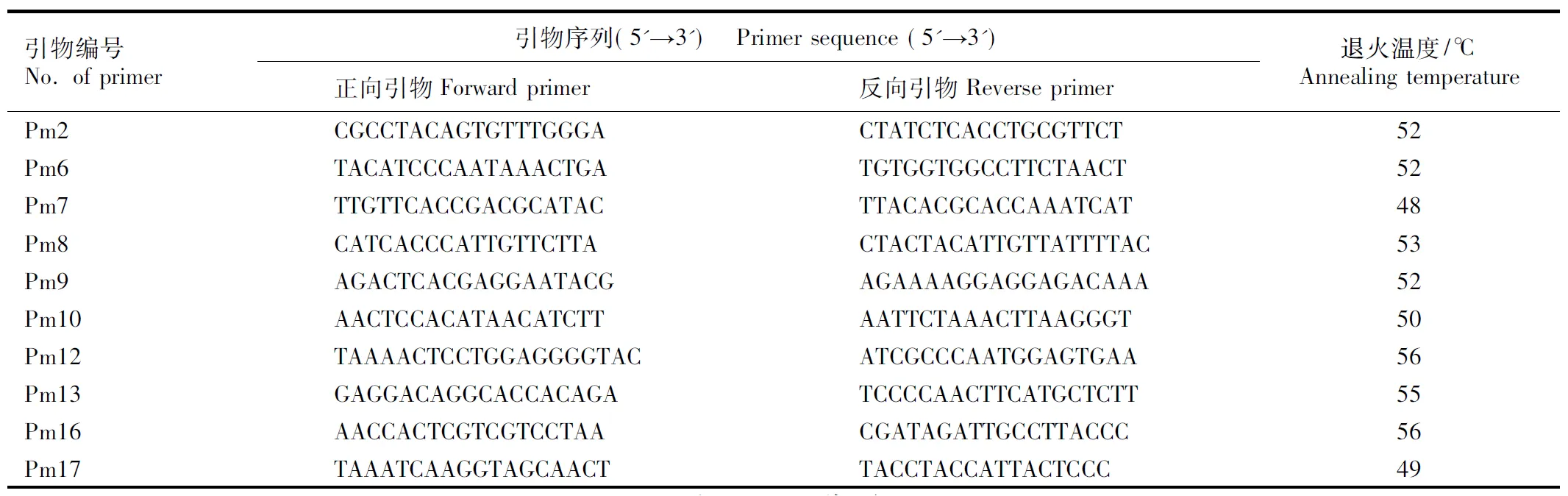
引物编号No. of primer引物序列(5'→3') Primer sequence (5'→3')正向引物Forward primer反向引物Reverse primer退火温度/℃Annealing temperaturePm2CGCCTACAGTGTTTGGGACTATCTCACCTGCGT-TCT52Pm6TACATCCCAATAAACTGATGTGGTGGCCTTCTA-ACT52Pm7TTGTTCACCGACGCATACTTACACGCACCAAAT-CAT48Pm8CATCACCCATTGTTCTTACTACTACATTGT-TATTTTAC53Pm9AGACTCACGAGGAATACGAGAAAAGGAGGAGA-CAAA52Pm10AACTCCACATAACATCTTAATTCTAAACTTA-AGGGT50Pm12TAAAACTCCTGGAGGGG-TACATCGCCCAATGGAGT-GAA56Pm13GAGGACAGGCACCACAGATCCCCAACTTCAT-GCTCTT55Pm16AACCACTCGTCGTCCTAACGATAGATTGCCTTAC-CC56Pm17TAAATCAAGGTAGCAACTTACCTACCATTACTCCC49
1.3 数据分析
使用Genepop 4.0软件检验位点间是否存在连锁不平衡及各居群是否偏离哈温平衡;使用FSTAT 2.9.3软件计算各居群的等位基因数、遗传分化系数和近交系数;使用Popgene 3.2软件计算各居群的观测杂合度和期望杂合度;使用GenAlEx 6软件计算各居群的有效等位基因数和稀有等位基因数。
使用TFPGA软件对居群间的遗传分化系数和遗传距离与地理距离的相关性进行Mantel检验;使用BayesAss 1.3软件计算居群间的基因流;使用Barrier 2.2软件分析居群间可能存在的主要基因流屏障;使用Arlequin 3.5软件对同型花和二型花居群间、所有居群间及各居群内的遗传变异进行分子方差分析(AMOVA);使用Structure 2.2软件对所有居群及样株进行聚类分析,K值设为6,重复分析8次;burn-in设为100 000,run-length设为1 000 000,使用混合模型(admixture model)[18]计算ΔK值。使用GenAlEx 6软件进行主坐标分析(PCoA)并制图。
使用SPSS 19.0统计分析软件对相关数据进行统计和分析。
2 结果和分析
2.1 安徽羽叶报春居群的遗传多样性分析
采用供试10对SSR引物对安徽羽叶报春6个居群叶片的总DNA进行PCR扩增,其中,引物Pm10对铜陵市义安区凤凰山(FHS)居群19个单株的扩增结果见图1。10对SSR引物在6个居群的117个单株中共扩增出129个等位基因,平均每对引物扩增出约13个等位基因。Genepop软件检测结果表明:仅少量位点在个别居群中存在连锁现象,未发现有位点在3个以上居群中与其他位点连锁。
对安徽羽叶报春各居群的遗传多样性分析结果(表3)表明:安徽羽叶报春6个居群的等位基因数、有效等位基因数和稀有等位基因数的平均值分别为4.8、2.8和2.7,观测杂合度、期望杂合度和近交系数的平均值分别为0.29、0.43和0.38。其中,FHS居群的等位基因数、有效等位基因数、稀有等位基因数、观测杂合度和期望杂合度均最低,分别为2.5、1.8、0.0、0.05和0.31,说明该居群的遗传多样性最低;池州市石台县立新村(LXC)和池州市石台县莲花田(LHT)居群的上述指标均较高,2个居群的等位基因数分别为6.4和7.5,有效等位基因数分别为3.2和4.7,稀有等位基因数分别为4.0和8.0,观测杂合度分别为0.48和0.37,期望杂合度分别为0.53和0.56,说明这2个居群的遗传多样性相对较高。值得注意的是,FHS居群的近交系数最高(0.81),池州市贵池区阴山(YS)居群的近交系数也较高(0.51),而LXC和LHT居群的近交系数相对较低(分别为0.11和0.36)。

1-19:单株编号 No. of individual;M:DNA marker.图1 引物Pm10对安徽羽叶报春铜陵市义安区凤凰山居群19个单株的扩增结果Fig. 1 Amplification result of nineteen individuals in population of Primula merrilliana Schltr. from Fenghuangshan in Yian District of Tongling City by primer Pm10
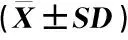
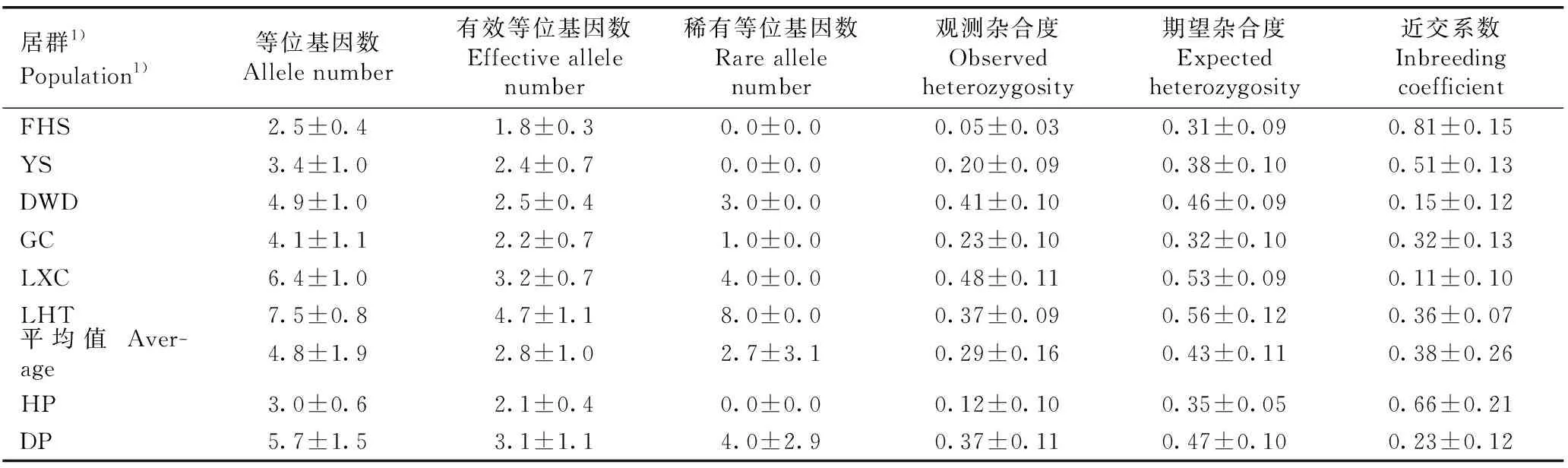
居群1)Population1)等位基因数Allele number有效等位基因数Effective allele number稀有等位基因数Rare allele number观测杂合度Observed heterozygosity期望杂合度Expected heterozygosity近交系数Inbreeding coefficientFHS2.5±0.41.8±0.30.0±0.00.05±0.030.31±0.090.81±0.15YS3.4±1.02.4±0.70.0±0.00.20±0.090.38±0.100.51±0.13DWD4.9±1.02.5±0.43.0±0.00.41±0.100.46±0.090.15±0.12GC4.1±1.12.2±0.71.0±0.00.23±0.100.32±0.100.32±0.13LXC6.4±1.03.2±0.74.0±0.00.48±0.110.53±0.090.11±0.10LHT7.5±0.84.7±1.18.0±0.00.37±0.090.56±0.120.36±0.07平均值 Aver-age4.8±1.92.8±1.02.7±3.10.29±0.160.43±0.110.38±0.26HP3.0±0.62.1±0.40.0±0.00.12±0.100.35±0.050.66±0.21DP5.7±1.53.1±1.14.0±2.90.37±0.110.47±0.100.23±0.12
1)FHS:铜陵市义安区凤凰山 Fenghuangshan in Yian District of Tongling City;YS:池州市贵池区阴山 Yinshan in Guichi District of Chizhou City;DWD:池州市石台县大王洞 Dawangdong in Shitai County of Chizhou City;GC:池州市贵池区谷村 Gucun in Guichi District of Chizhou City;LXC:池州市石台县立新村 Lixincun in Shitai County of Chizhou City;LHT:池州市石台县莲花田 Lianhuatian in Shitai County of Chizhou City. HP:同型花居群 Homostyled population;DP:二型花居群 Distyled population.
对安徽羽叶报春同型花和二型花居群的遗传多样性分析结果(表3)表明:同型花居群的等位基因数、有效等位基因数、观测杂合度和期望杂合度(分别为3.0、2.1、0.12和0.35)均低于二型花居群(分别为5.7、3.1、0.37和0.47),说明安徽羽叶报春同型花居群的遗传多样性低于二型花居群。然而,同型花居群(0.66)的近交系数却明显高于二型花居群(0.23)。值得注意的是,所有稀有等位基因均出现在二型花居群中,而在同型花居群中未检测到稀有等位基因。
2.2 安徽羽叶报春居群的遗传结构分析
安徽羽叶报春6个居群间的遗传分化系数和遗传距离见表4。由表4可以看出:安徽羽叶报春6个居群间遗传分化系数的平均值为0.34,其中,铜陵市义安区凤凰山(FHS)和池州市贵池区阴山(YS)居群间的遗传分化系数最大(0.45),池州市石台县大王洞(DWD)和池州市石台县立新村(LXC)居群间的遗传分化系数最小(0.22)。
由表4还可以看出:安徽羽叶报春6个居群间遗传距离的平均值为0.56,其中,FHS和YS居群间的遗传距离最大(0.74),FHS和LXC居群间的遗传距离最小(0.37)。
Mantel检验结果表明:安徽羽叶报春各居群间的遗传分化系数和遗传距离与地理距离的相关系数分别为0.356(P=0.193)和0.250(P=0.370),说明安徽羽叶报春各居群间的遗传分化系数和遗传距离与地理距离无相关性。
安徽羽叶报春6个居群间的基因流计算结果(表5)表明:由LXC和池州市石台县莲花田(LHT)居群向其他居群的基因流较大,为0.057~0.065;由DWD和池州市贵池区谷村(GC)居群向其他居群的基因流为0.050~0.053;由FHS和YS居群向其他居群的基因流较小,为0.044~0.046。上述结果说明越靠近秋浦河流域上游的安徽羽叶报春居群,其基因更容易向其他居群流动。进一步分析结果表明:第1基因流屏障介于FHS和其他居群间,第2基因流屏障介于二型花和同型花居群间。
表4安徽羽叶报春6个居群间的遗传分化系数和遗传距离1)
Table4GeneticdifferentiationcoefficientandgeneticdistanceamongsixpopulationsofPrimulamerrillianaSchltr.1)
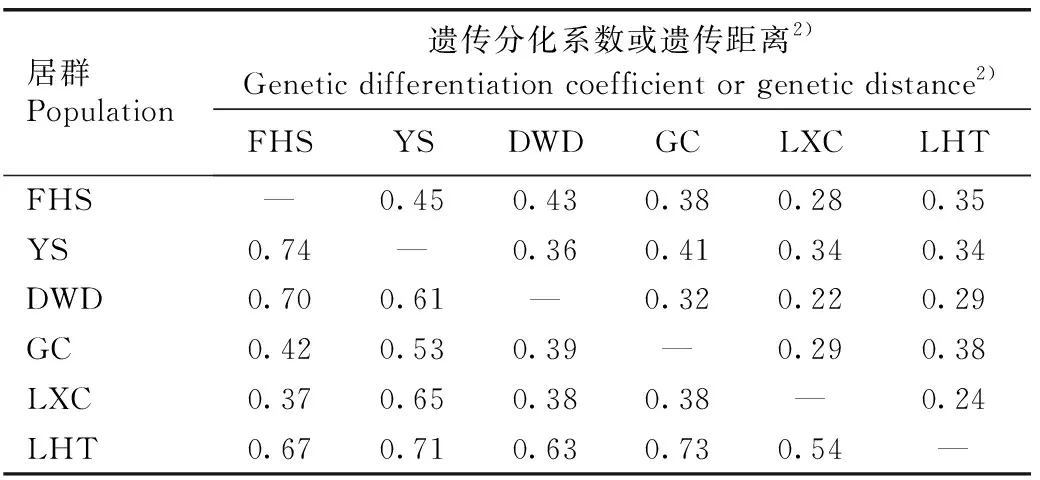
居群Population遗传分化系数或遗传距离2)Genetic differentiation coefficient or genetic distance2)FHSYSDWDGCLXCLHTFHS—0.450.430.380.280.35YS0.74—0.360.410.340.34DWD0.700.61—0.320.220.29GC0.420.530.39—0.290.38LXC0.370.650.380.38—0.24LHT0.670.710.630.730.54—
1)FHS:铜陵市义安区凤凰山 Fenghuangshan in Yian District of Tongling City;YS:池州市贵池区阴山 Yinshan in Guichi District of Chizhou City;DWD:池州市石台县大王洞 Dawangdong in Shitai County of Chizhou City;GC:池州市贵池区谷村 Gucun in Guichi District of Chizhou City;LXC:池州市石台县立新村 Lixincun in Shitai County of Chizhou City;LHT:池州市石台县莲花田 Lianhuatian in Shitai County of Chizhou City.
2)“—”上方和下方数据分别为遗传分化系数和遗传距离 Data above and below “—” are genetic differentiation coefficient and genetic distance,respectively.
表5安徽羽叶报春6个居群间的基因流1)
Table5GeneflowamongsixpopulationsofPrimulamerrillianaSchltr.1)
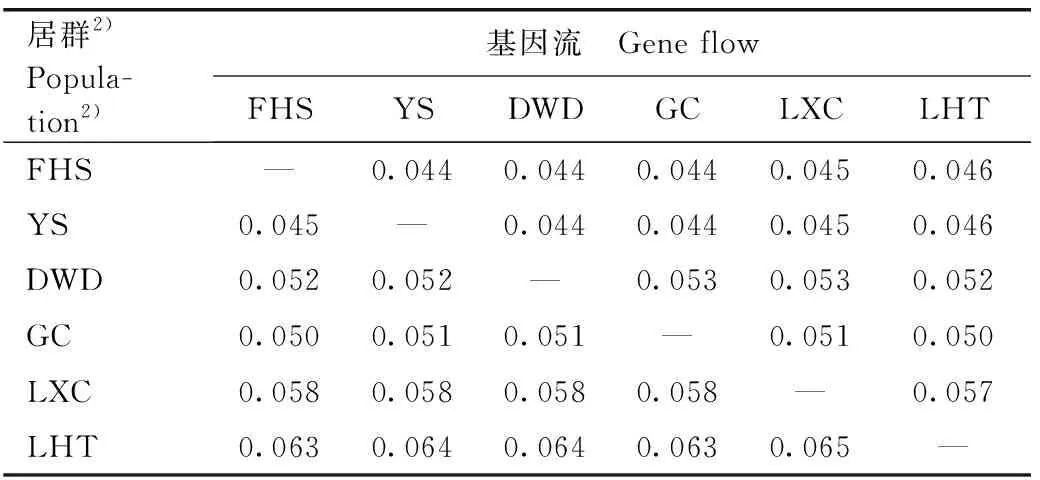
居群2)Popula-tion2)基因流 Gene flowFHSYSDWDGCLXCLHTFHS—0.0440.0440.0440.0450.046YS0.045—0.0440.0440.0450.046DWD0.0520.052—0.0530.0530.052GC0.0500.0510.051—0.0510.050LXC0.0580.0580.0580.058—0.057LHT0.0630.0640.0640.0630.065—
1)FHS:铜陵市义安区凤凰山 Fenghuangshan in Yian District of Tongling City;YS:池州市贵池区阴山 Yinshan in Guichi District of Chizhou City;DWD:池州市石台县大王洞 Dawangdong in Shitai County of Chizhou City;GC:池州市贵池区谷村 Gucun in Guichi District of Chizhou City;LXC:池州市石台县立新村 Lixincun in Shitai County of Chizhou City;LHT:池州市石台县莲花田 Lianhuatian in Shitai County of Chizhou City.
2)表中数据为由此列中居群向其他居群的基因流 Data in the table are gene flows from populations in this column to other populations.
分子方差分析(AMOVA)结果(表6)表明:同型花和二型花居群间的遗传差异不显著(P>0.05);各居群间出现一定的遗传分化,居群间遗传变异占总遗传变异的36.40%(P<0.001);64.76%的遗传变异存在于居群内,说明安徽羽叶报春居群的主要遗传变异存在于居群内。
聚类分析结果表明:lnP(D)值随着K值的升高而持续且均匀的上升,且ΔK值无明显峰值(最大值仅为6.75),表明安徽羽叶报春各居群间的聚类结果并不明显。
主坐标分析(PCoA)结果(图2)表明:2个同型花居群的供试植株总体上均位于4个二型花居群供试植株的外围,并且不同居群间的供试植株均存在一定程度的分离,但是同型花和二型花居群间的分离程度并未明显大于二型花居群间的分离程度,说明安徽羽叶报春的同型花和二型花植株间并没有产生明显的遗传分化。
表6安徽羽叶报春6个居群的分子方差分析(AMOVA)
Table6Analysisofmolecularvariance(AMOVA)amongsixpopulationsofPrimulamerrillianaSchltr.
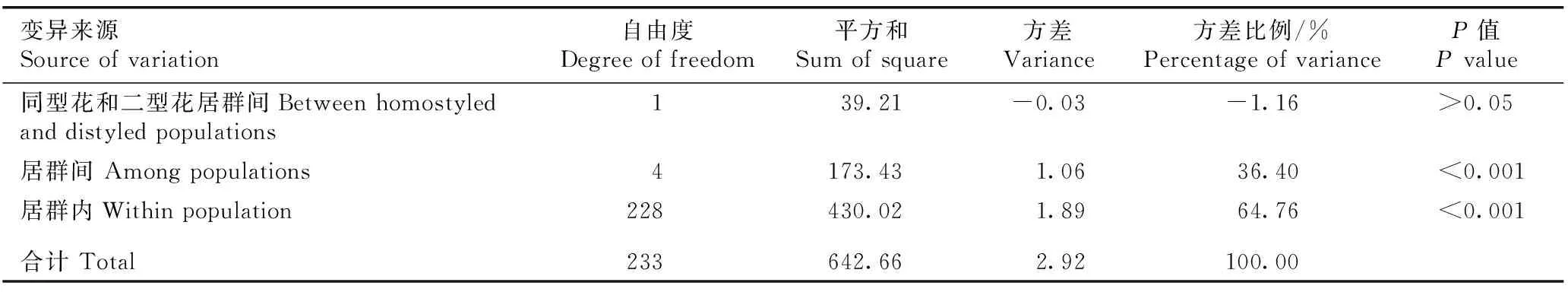
变异来源Source of variation自由度Degree of freedom平方和Sum of square方差Variance方差比例/%Percentage of varianceP值P value同型花和二型花居群间Between homostyled and distyled populations139.21-0.03-1.16>0.05居群间 Among populations4173.431.0636.40<0.001居群内Within population228430.021.8964.76<0.001合计 Total233642.662.92100.00
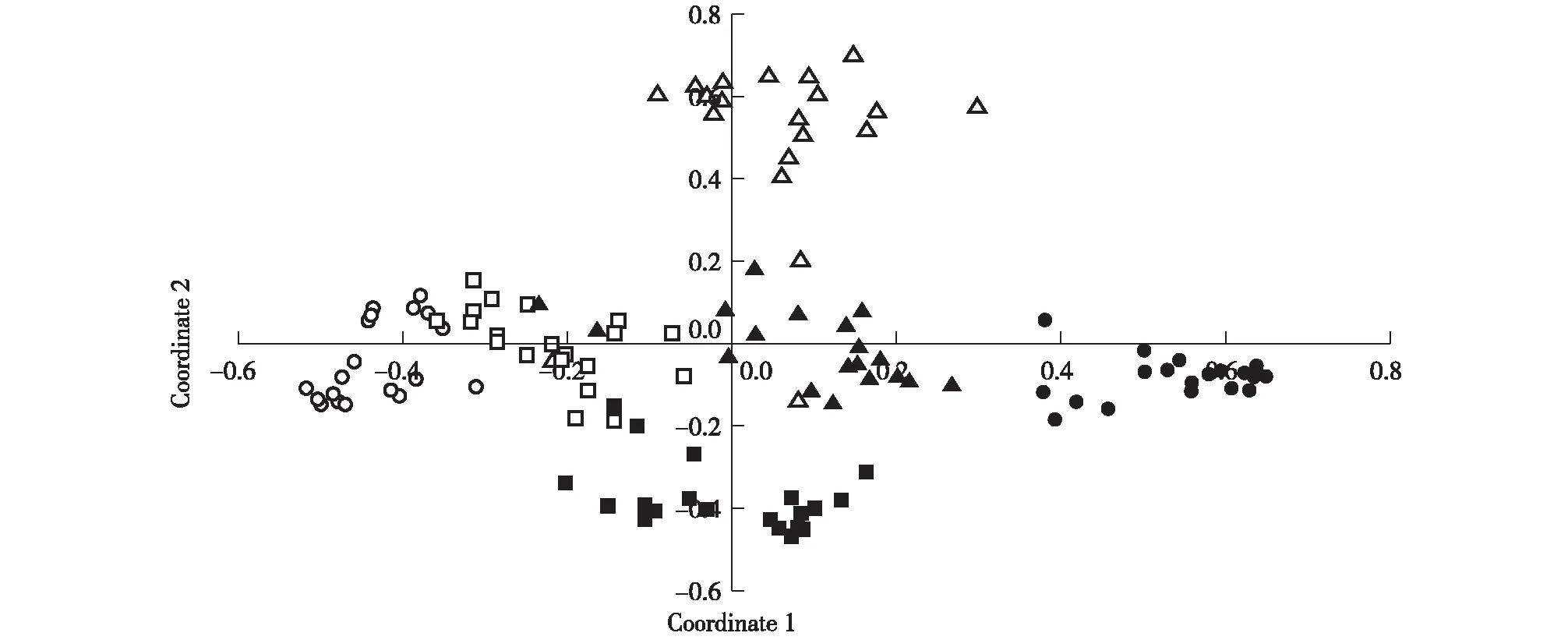
●:铜陵市义安区凤凰山Fenghuangshan in Yian District of Tongling City;○:池州市贵池区阴山 Yinshan in Guichi District of Chizhou City;■:池州市贵池区谷村 Gucun in Guichi District of Chizhou City;□:池州市石台县大王洞 Dawangdong in Shitai County of Chizhou City;▲:池州市石台县立新村 Lixincun in Shitai County of Chizhou City;△:池州市石台县莲花田 Lianhuatian in Shitai County of Chizhou City.图2 安徽羽叶报春6个居群的主坐标分析(PCoA)Fig. 2 Principal coordinate analysis (PCoA) on six populations of Primula merrilliana Schltr.
3 讨 论
安徽羽叶报春为中国特有的野生珍稀花卉种类,具有较高的园艺开发和应用价值[19],现仅分布于安徽省南部的丘陵山区。本研究涉及的6个自然居群均位于安徽羽叶报春分布区的西北角,即牯牛降北坡及九华山西北坡的秋浦河流域。研究结果显示:安徽羽叶报春6个居群的等位基因数、有效等位基因数、稀有等位基因数、观测杂合度和期望杂合度的平均值均较低,分别为4.8、2.8、2.7、0.29和0.43,说明供试安徽羽叶报春各居群的遗传多样性较低;并且,4个二型花居群的等位基因数、观测杂合度和期望杂合度的平均值分别为5.7、0.37和0.47,明显低于安徽羽叶报春分布区中心的黄山及其周边区域的居群(等位基因数、观测杂合度和期望杂合度的平均值分别为8.1、0.46和0.59)[15],据此判断供试安徽羽叶报春居群遗传多样性的分布基本符合“核心-边缘”假说[20]。与中心分布区居群相比,由于安徽羽叶报春边缘分布区居群对生境的适应性较差,居群规模受气候因子变化的影响较大,更易受到瓶颈效应的影响。另外,由于安徽羽叶报春边缘分布区居群通常缺少传粉昆虫,其自交繁育系统更易被保留,居群的近交系数明显升高。综上所述,多因子综合作用促使安徽羽叶报春边缘分布区居群的遗传多样性明显下降。
本研究中,铜陵市义安区凤凰山(FHS)和池州市贵池区阴山(YS)居群为同型花居群,位于安徽羽叶报春分布区的最北缘,其等位基因数、观测杂合度和期望杂合度的平均值分别为3.0、0.12和0.35,明显高于安徽羽叶报春的近缘种堇叶报春(PrimulacicutariifoliaPax)(等位基因数、观测杂合度和期望杂合度的平均值分别为1.4、0.01和0.07)[21],但却低于供试二型花居群。Mast等[8]认为,报春花属中约10%的同型花种类由二型花种类进化而来,花型进化趋势为由二型花向同型花转变,据此推断FHS和YS居群也是由二型花植株演化而来,可能是由于基因重组等原因在二型花居群中出现了少量的同型花植株,这些同型花植株的雌雄蕊等高且自交亲和性强,无需传粉昆虫便可采用自交方式完成有性生殖过程,并且,这些同型花植株具有较好的生殖保障和自动选择优势[22],在分布区边缘居群中具有较高的适合度,从而被保留,最终形成同型花居群。植物的繁育系统是影响其居群遗传多样性的重要因子,在由异交向自交的转变过程中常常伴随着有效种群减小、连锁不平衡加剧及基因清除效应增强等现象,进而导致居群的遗传多样性降低甚至消失[23]。相关研究结果表明:具有自交繁育系统的植物种类的遗传多样性明显低于具有异交繁育系统的近缘种[24-25],即使是同种类的不同居群,也会由于繁育系统的自交化转变而导致其遗传多样性明显下降[23,26-27],这可能是导致安徽羽叶报春FHS和YS 2个同型花居群遗传多样性低于其附近区域二型花居群的主要原因。
本研究中,秋浦河流域上游的池州市石台县莲花田(LHT)和池州市石台县立新村(LXC)居群的遗传多样性均较高,且由这2个居群向其他居群的基因流较大,说明研究区域内的安徽羽叶报春居群的遗传多样性与居群位置有一定关系,建议加强对秋浦河流域上游安徽羽叶报春居群的保护,尤其应注意对居群生境的保护。位于秋浦河流域下游低矮丘陵山区的FHS和YS居群为同型花居群,其繁育系统已经变为自交繁育,与二型花居群的繁育系统(异交繁育)已经明显不同,且与二型花居群间存在明显的基因流屏障,因此,建议将其视为不同于二型花居群的独立单元来进行遗传保护。
参考文献:
[1] BARRETT S C H,JESSON L K,BAKER A M. The evolution and function of stylar polymorphisms in flowering plants[J]. Annals of Botany,2000,85(Supplement A):253-265.
[2] NOWAK M D,RUSSO G,SCHLAPBACH R,et al. The draft genome ofPrimulaverisyields insights into the molecular basis of heterostyly[J]. Genome Biology,2015,16(1):12.
[3] DARWIN C. On the two forms,or dimorphic condition,in the species ofPrimula,and on their remarkable sexual relations[J]. Botanical Journal of the Linnean Society,1862,6(22):77-96.
[4] BARRETT,S C H,SHORE J S. New insights on heterostyly:comparative biology,ecology and genetics[M]∥FRANKLIN-TONG V E. Self-incompatibility in Flowering Plants:Evolution,Diversity,and Mechanisms. Berlin:Springer-Verlag,2008:3-32.
[5] PIPER J G,CHARLESWORTH B,CHARLESWORTH D. Breeding system evolution inPrimulavulgarisand the role of reproductive assurance[J]. Heredity,1986,56(2):207-217.
[6] ECKERT C G,KALISZ S,GEBER M A,et al. Plant mating systems in a changing world[J]. Trends in Ecology and Evolution,2010,25(1):35-43.
[7] KELSO S. The genusPrimulaas a model for evolution in the Alaskan flora[J]. Arctic and Alpine Research,1992,24(1):82-87.
[8] MAST A R,KELSO S,CONTI E. Are any primroses (Primula) primitively monomorphic?[J]. New Phytologist,2006,171(3):605-616.
[9] 中国科学院中国植物志编辑委员会. 中国植物志:第五十九卷第二分册[M]. 北京:科学出版社,1990:55-57.
[10] RICHARDS J. Primula[M]. Portland:Timber Press,2003:1-69.
[11] 张小平,陈明林. 安徽羽叶报春与毛茛叶报春的遗传多样性研究与新资源评估[J]. 植物资源与环境学报,2003,12(3):1-5.
[12] 杜丹丹,邵剑文. 中国特有濒危植物安徽羽叶报春的研究现状及展望[J]. 安徽师范大学学报(自然科学版),2010,33(6):562-565.
[13] 陈明林,张小平,李晓红. 安徽羽叶报春和毛茛叶报春的微形态特征[J]. 植物资源与环境学报,2004,13(3):18-24.
[14] 陈明林,张小平,孟凡庭. 皖南奇葩:安徽羽叶报春[J]. 植物杂志,2001(2):6-7.
[15] SHAO J W,WANG J,XU Y N,et al. Genetic diversity and gene flow within and between two different habitats ofPrimulamerrilliana(Primulaceae),an endangered distylous forest herb in eastern China[J]. Botanical Journal of the Linnean Society,2015,179(1):172-189.
[16] SHAO J W,CHEN W L,PENG Y Q,et al. Genetic diversity within and among populations of the endangered and endemic speciesPrimulamerrillianain China[J]. Biochemical Systematics and Ecology,2009,37(6):699-706.
[17] DOYLE J. DNA protocols for plants-CTAB total DNA isolation[M]∥HEWITT G M,JOHNSTON A W B,YOUNG J P W. Molecular Techniques in Taxonomy. Berlin:Springer-Verlag,1991:283-293.
[18] EVANNO G,REGNAUT S,GOUDET J. Detecting the number of clusters of individuals using the software STRUCTURE:a simulation study[J]. Molecular Ecology,2005,14(8):2611-2620.
[19] 邵剑文,张小平,张中信,等. 安徽羽叶报春的有效传粉昆虫及花朵密度和种群大小对传粉效果的影响[J]. 植物分类学报,2008,46(4):537-544.
[20] MAYR E. Animal Species and Evolution[M]. Cambridge:Harvard University Press,1963:226-228.
[21] WANG D Y,CHEN Y J,ZHU H M,et al. Highly differentiated populations of the narrow endemic and endangered speciesPrimulacicutariifoliain China,revealed by ISSR and SSR[J]. Biochemical Systematics and Ecology,2014,53(8):59-68.
[22] 张大勇. 植物生活史进化与繁殖生态学[M]. 北京:科学出版社,2004:319-320.
[24] KOELLING V A,HAMRICK J L,MAURICIO R. Genetic diversity and structure in two species ofLeavenworthiawith self-incompatible and self-compatible populations[J]. Heredity,2011,106(2):310-318.
[25] NYBOM H,BARTISH I V. Effects of life history traits and sampling strategies on genetic diversity estimates obtained with RAPD markers in plants[J]. Perspectives in Plant Ecology,Evolution and Systematics,2000,3(2):93-114.
[26] HERLIHY C R,ECKERT C G. Genetic cost of reproductive assurance in a self-fertilizing plant[J]. Nature,2002,416(6878):320-323.
[27] MABLE B K,ADAM A. Patterns of genetic diversity in outcrossing and selfing populations ofArabidopsislyrata[J]. Molecular Ecology,2007,16(17):3565-3580.
猜你喜欢
植物研究(2022年5期)2022-10-12
河北科技师范学院学报(2022年2期)2022-08-26
北极光(2022年1期)2022-02-23
浙江中医药大学学报(2021年6期)2021-07-12
军工文化(2021年2期)2021-03-30
中国粮油学报(2020年12期)2021-01-09
幽默大师(漫话国学)(2020年9期)2020-09-15
活力(2019年15期)2019-09-25
航空模型(2017年2期)2017-05-22
中国非金属矿工业导刊(2015年5期)2015-12-22
