
荷花由单瓣到千瓣的演化及调控机制探讨*
2018-06-25 08:39秦密刘凤栾张大生田代科
自然杂志 2018年3期
秦密,刘凤栾,张大生,田代科†
①上海辰山植物园 中国科学院上海辰山植物科学研究中心,上海 201602;②上海师范大学 生命与环境科学学院,上海 200234
“出淤泥而不染,濯清涟而不妖”,精炼地总结出荷花的生活习性及特点;“接天莲叶无穷碧,映日荷花别样红”,生动地描绘出荷花景色之美;“青荷盖绿水,芙蓉披红鲜。下有并根藕,上有并头莲”则借独特稀奇的“并蒂莲”现象暗喻了美好爱情。从古至今,之所以有如此之多的诗句和文章描绘、赞美荷花,与其集食用、药用、观赏多种用途于一身,并不断被赋予“纯洁、清正、高雅”等人格象征紧密关联。因此,在经济需求(多种实用价值)及精神需求(象征意义)的双重选择下,各种各样的荷花品种不断被人们培育出来。迄今为止,全球仅观赏荷花品种就已达2 000多个。就花的被片数而言,观赏荷花品种已演化出单瓣、重瓣、重台及千瓣等多种类型[1]。在园林园艺应用中,个体花期较长、造型丰富多变的重瓣荷花(如‘友谊牡丹’‘中山红台’‘至尊千瓣’和‘光岳楼’)观赏价值更高,造景效果更好。事实上,野生荷花都是单瓣类型,花瓣数量一般24~28片,经过长时间的自然和人为选择之后,才出现了重瓣及千瓣荷花。那么,荷花的单瓣、重瓣是如何划分的?区别在何处?花瓣重瓣化现象又是如何发生的呢?本文就此展开论述。
莲科植物最早的化石发现于早白垩世劳亚古陆中纬度地区,广布于亚洲大陆和北美洲。现仅有莲属(Nelumbo)2个种,即分布于亚洲和澳大利亚北部的亚洲莲(Nelumbo nuceferaGaertn.)和中北美洲的美洲莲(N. luteaWilld.)[2]。作为荷花的世界分布中心和栽培中心之一[3],我国拥有世界上最丰富的荷花种质资源,除了争奇斗艳、丰富多样的栽培品种,还包括许多野生类型:黑龙江流域的野生莲,黄河长江流域的‘白洋淀红’莲、‘微山湖红’莲和‘洪湖红’莲等,表现为单瓣、红色或粉红色等表型。19世纪末,在大连市普兰店发掘的古莲子,培育成苗后开花亦为单瓣红色(图1)。我们通过分子标记检测发现,它与已发现的‘古代’莲和黑龙江等东北野生莲的亲缘关系最近,暗示了其野生莲身份(待发表)。

图1 盛开的‘普兰店古’莲
1 ‘普兰店古’莲
19世纪末,辽宁大连普兰店的居民从当地旧河床中发现古莲子。1923年,日本植物学博士大贺一郎(Ichiro Ohga)首次将普兰店出土的古莲子成功培育成植株,并根据发现古莲子处的河床岩石、泥土等地质学特征推测这些古莲子的年龄在400年以上[4]。此后,关于‘普兰店古’莲年龄的报道相继出现。1951年美国著名放射化学家利比(Libby)用放射性碳技术测出‘普兰店古’莲的年龄为1 040±210年[5];1975年,经中国科学院考古研究所测定1952年北京植物园在普兰店收集的古莲子寿命为915±80年[7];1982年荷兰科学家普利斯特列(Priestley)等用放射性同位素测定一粒‘普兰店古’莲种子的种龄约为466年[6]。1955年中国科学院植物研究所成功地将普兰店的古莲子培育开花,引起很大轰动[7]。古莲子之所以历经千年能保存下来并发芽开花,同其致密的果皮结构、莲子含抗氧化物质以及莲子埋藏在地下密闭环境等条件密不可分。得益于普兰店的爱莲人徐刚等对古莲的积极保护,如今普兰店已成为中国最大的原生态古莲基因库基地。2008年,徐刚还将‘普兰店古’莲所结莲子搭载“神舟七号”载人飞船遨游太空,返回后育出了‘飞天古’莲等优良品种。
2 荷花品种瓣型演化及分类
在长久的自然和人工选择下,观赏荷花出现单瓣、半重瓣、重瓣、重台和千瓣等多种类型,极大地提升了观赏价值。其中,单瓣是指花的各部分发育正常,花被形态完整,无雌雄蕊发生瓣化,花被片数常在30以下。自然界的野生莲‘微山湖红’‘白洋淀红’和千年 ‘普兰店古’莲(图1、图2a)以及传统品种‘单洒锦’(图2A)等都属于单瓣类型。半重瓣:仅有少部分雄蕊发生瓣化,最内层的一些被片可观察到雄蕊瓣化的痕迹或过渡类型,花被片数小于单瓣的2倍,常在30~60。‘红衣舞女’(图2b)和‘霞披’(图2B)等均为此类。重瓣:花的雄蕊多数已瓣化,雌蕊部分或全部转变为花瓣或其类似结构,被片总数通常在60枚以上,为单瓣被片数目的2倍多,如‘大洒锦’(图2c)和‘友谊牡丹’等(图2C)。重台:为高度半重瓣或重瓣花的特殊类型,部分雄蕊保留,心皮明显泡化或特化成花瓣状结构,形成上下双层花被结构,但花托完整或出现退化,‘光岳楼’(图2d)和‘红台’莲(图2D)为其代表。千瓣:实为重瓣花的极端类型,也属于全重瓣,‘至尊千瓣’(图2e)、‘千瓣’莲(图2E)为此类型,其中后者不仅外部被片数大量增加,一朵花内还形成多个发育中心,包含2~5个花心,每个花心由若干细小花瓣组成。需注意的是,上述各类型间的花瓣数目并不存在明确分界[8-9]。

图2 不同花型的荷花品种:单瓣(a‘普兰店古’莲、A‘单洒锦’)、半重瓣(b‘红衣舞女’、B‘霞披’)、重瓣(c‘大洒锦’、C‘友谊牡丹’)、重台(d‘光岳楼’、D‘中山红台’)和千瓣(e‘至尊千瓣’、E‘千瓣’莲)
3 重瓣荷花的来源
荷花属于同被花,花萼与花瓣没有明确的分界。重瓣荷花的瓣化程度或花被数量差别很大,例如‘大洒锦’平均为93,‘中山红台’平均为306,而‘千瓣’莲的被片数最多,上千至几千(图3)。通过对这些重瓣类型花朵的形态解剖观察比较,认为其瓣化发生大致源于4种情况:雌蕊正常但雄蕊部分瓣化;雌蕊和雄蕊均部分瓣化;雌雄蕊全部瓣化以及花瓣的自然增加。具体描述如下:①雌蕊正常但雄蕊部分瓣化。此类多数为雄蕊向心式瓣化,即外围雄蕊转变为花瓣,而接近花托的雄蕊保持不变(图3A1),部分品种可观察到雄蕊发生瓣化的过渡形态——花瓣顶部有花药或附属物的残留(图3A2),即单个雄蕊瓣化的方向为花丝→花药→附属物(图3A3)。‘恋夏’‘友谊牡丹’和‘小羚羊’等属于此类型。②雌蕊和雄蕊均部分瓣化。随着雄蕊瓣化程度增加,雌蕊也发生瓣化(图3B1和B2),雌蕊变泡状或沿着腹缝线一侧开裂形成管状花瓣(图3 B3和B4),从而形成重瓣或重台型,但仍有一定数量的正常雄蕊存在(图3B2)。代表型品种如‘红台’莲和‘中山红台’等。③雌雄蕊全部瓣化。雌蕊、雄蕊几乎全部发生瓣化(图3 C1~C4),使得一朵花的被片数急剧增加,如‘光岳楼’和‘至尊千瓣’等。④花瓣的自然增加。‘千瓣’莲(图3 D1和D2)花蕾内部生长点不形成雄蕊和雌蕊原基,而是产生多个类似花瓣原基(花瓣分生增值中心)结构,常形成多个“花头”(图3D3),出现紧密相连的“花中花”景观,这些花头由细小花瓣密集组成(图3D4和D5),总花瓣数超过1 000,甚至多达数千。已有研究认为,荷花重瓣化的发生主要为前3种情况,即重瓣的花被片主要来源于雄蕊的向心式瓣化,其次是雌蕊瓣化,属于植物重瓣花起源6种方式(积累起源、苞片起源、雌雄蕊起源、台阁起源、重复起源和花序起源)中的雌雄蕊起源方式[10]。荷花重瓣化的第四种情况属于哪一种重瓣花起源方式目前尚未定论,是否存在积累起源或者是其他的起源还有待研究。
4 荷花重瓣化机制的研究进展
重瓣荷花品种的观赏价值很高,是荷花新品种育种的重要亲本之一,也是研究高等植物花朵重瓣化演化的适宜材料。然而,关于荷花重瓣化机制的研究报道很少。在细胞水平上,姜莉等[11]通过解剖重瓣‘友谊牡丹’莲和重台的‘红台’莲的花芽分化过程,发现重瓣和重台荷花均存在雌雄蕊的瓣化,其中重台荷花的雌雄蕊瓣化程度较大,并失去了性细胞的分化。针对单瓣、重瓣、重台、全重瓣及千瓣等5种花型荷花的花芽分化进行解剖分析,黄秀等[10]证明荷花重瓣化花瓣主要来源于雄蕊的向心式瓣化,其次是雌蕊瓣化,属于雌雄蕊起源。其中,全重瓣品种中有雌雄蕊原基的分化,但发育后期均转化为花瓣,其雄蕊原基转向花瓣的发育过程与牡丹和芍药雄蕊筒状花瓣的发育过程相似,大致也可分为雄蕊原基分化、转化发育、小花瓣形成和成熟等4个阶段。

图3 不同瓣化程度荷花的解剖结构:A1部分雄蕊瓣化的花朵,A2雄蕊瓣化被片残留花药或附属物痕迹,A3雄蕊及其瓣化过渡态;B1雌蕊发生泡化的花朵,B2雌蕊泡化的花托,B3花托及瓣化雌蕊解剖,B4 瓣化雌蕊;C1雄蕊全部瓣化、雌蕊全部泡化的‘光岳楼’,C2‘光岳楼’花朵的纵切,C3花托及泡化的心皮,C4完全泡化的心皮;D1‘千瓣’莲,D2一朵花中的一个“花头”,D3‘千瓣’莲的花心,D4‘千瓣’莲的退化花托,D5‘千瓣’莲多个花心的花托(D4放大图)
著名荷花研究及育种者黄国振认为,荷花的重瓣是隐性性状[12]:通过单瓣‘白花建’莲和重瓣红花‘红千叶’的杂交试验,F1代的所有杂合植株均表现为单瓣红花;对F1代进行人工自花授粉,结果发现F2代中分离出重瓣红花、单瓣红花和单瓣白花等各种不同的类型。此外,在比较了不同重瓣程度品种染色体组型结构后,他还认为:荷花“重瓣化”主要是由于某些染色体结构发生突变导致的,特别是高度重瓣化的‘重台’莲和‘并蒂’莲,其染色体结构的变异更为显著[13]。近年来,随着基因测序技术的迅猛发展,转录组测序分析成为研究植物花发育的一个新途径。我们课题组已利用二代测序技术对单瓣、重瓣和千瓣的荷花花蕾进行转录组测序,获得46 584个独立基因(unigene),涉及花器官发育、花色途径、糖脂合成与代谢,以及信号传导等相关基因信息[14],为荷花的花器官发育机制研究提供了丰富的分子水平信息。
植物外在形态由自身遗传物质和环境条件共同决定,荷花被片数增加由相关基因调控。1991年Coen等[15]通过对拟南芥(Arabidopsis thaliana)和金鱼草(Amtirrhinum majus)花器官的研究,提出了调控植物花器官发育的经典ABC模型。该模型认为:A类基因单独决定萼片特性,A和B类基因共同决定花瓣特性,B和C类基因共同决定雄蕊特性,C类基因单独控制心皮特性。也就是说,一类基因控制相邻两轮花器官的发育:萼片(A)、花瓣(A+B)、雄蕊(B+C)、心皮(C)。随后,在研究矮牵牛(Petunia hybrida)的胚珠发育调控时,克隆到仅在胚珠原基、珠被和珠柄中表达的FBP11基因,并将其命名为D类基因[16],结合后续发现的协同调控萼片发育的E类基因,经典ABC模型被发展为ABCDE模型:萼片(A+E)、花瓣(A+B+E)、雄蕊(B+C+E)、心皮(C+E)、胚珠(D+E)(图4)。其中,B类和C类基因在高等植物重瓣花发育中起着重要作用,它们的突变或异常表达可诱导花朵的重瓣化。例如:金鱼草C功能AG同源基因FAR的缺失突变导致雄蕊瓣化形成重瓣花;麝香百合因其C类基因向中央收缩使外轮雄蕊变成花瓣而形成重瓣花[17]。目前荷花花发育的具体调控机制尚未清晰,因荷花重瓣化主要源于其雌雄蕊群,所以我们推测,调控荷花重瓣化演化的关键基因应属于B类和C类。
随着生物技术的不断发展,研究者对荷花花发育调控机制的了解逐渐深入。转基因体系的不断完善,可以对荷花开展分子辅助育种及更有针对性的分子育种,培育出更多新奇、独特的重瓣荷花品种,丰富和提高荷花的观赏性,进而提升水域景观及园林造景的观赏效果。
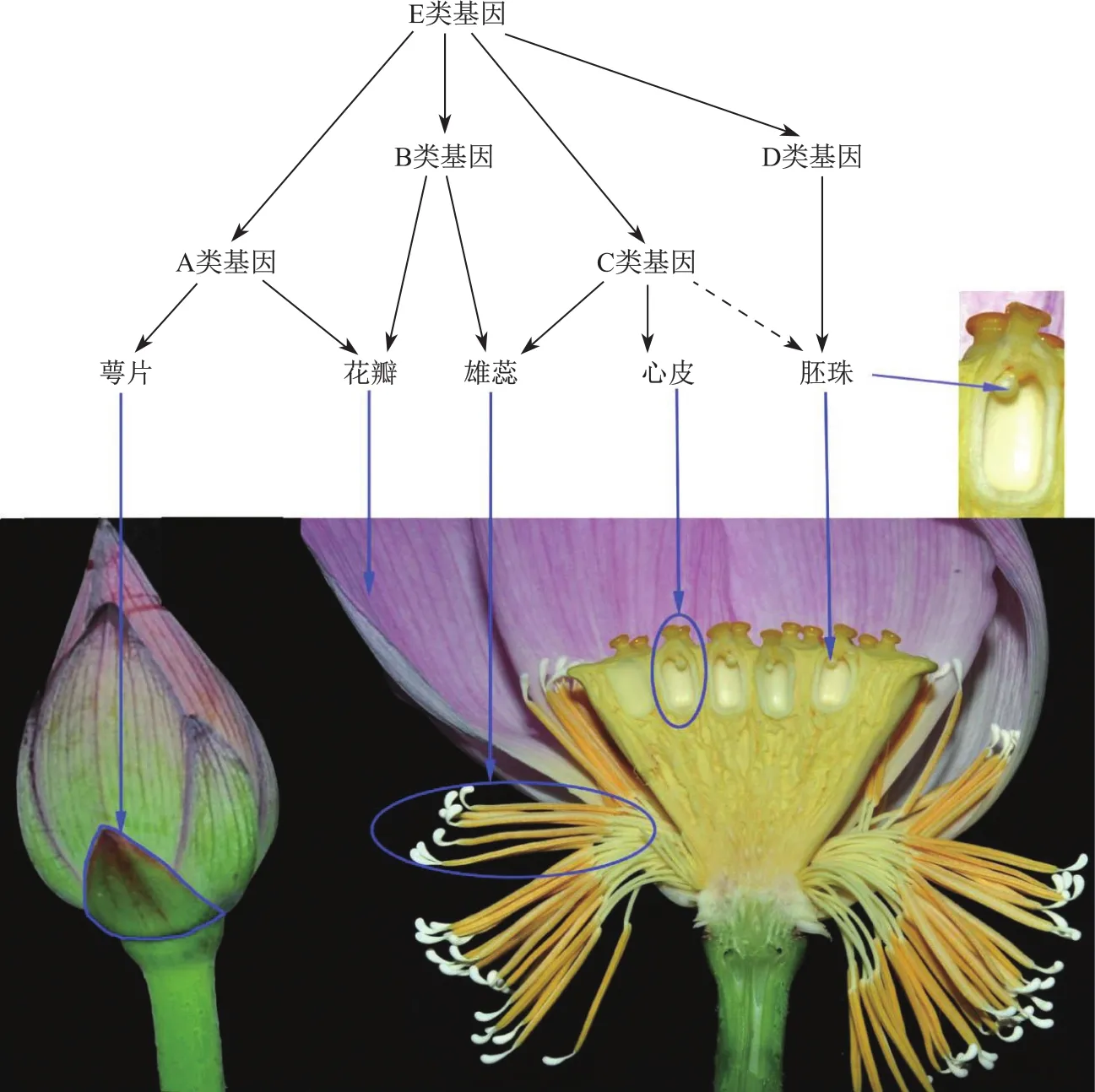
图4 植物花器官发育的ABCDE模型(参考Kim等[18])。A+E类基因调控萼片发育,A+B+E类基因调控花瓣发育,B+C+E类基因调控雄蕊发育,C+E类基因调控心皮发育,D+E类基因调控胚珠发育,C类基因也可能参与了调控胚珠发育
(2017年10月9日收稿)■
[1]田代科, 张大生. 莲叶何田田——世界荷花研究进展[J]. 生命世界,2014, 6: 40-45.
[2]LI Y, SVETLANA P, YAO J X, et al. A review on the taxonomic,evolutionary and phytogeographic studies of the lotus plant(Nelumbonaceae:Nelumbo) [J]. Acta Geologica Sinica, 2014, 88(4):1252-1261.
[3]王其超, 张行言. 中国荷花品种图志[M]. 北京: 中国林业出版社,2005: 2.
[4]OHGA I. On the longevity of seeds ofNelumbo nucifera[J]. 植物学杂志, 1923, 37: 439-444.
[5]LIBBY W F. Radiocarbon dates II [J]. Science, 1951, 114: 291-296.
[6]薛建华, 徐刚. 普兰店古莲[J]. 生命世界, 2015, 6: 20-25.
[7]张义君. 千年古莲苏醒了[J]. 化石, 1978, 1: 22-23.
[8]倪学明. 莲的品种分类研究[J]. 园艺学报, 1983, 10(3): 207-210.
[9]王其超, 张行言, 胡春根. 荷花品种分类新系统[J]. 植物科学学报,1997, 15(1): 19-26.
[10]黄秀, 田代科, 张微微, 等. 荷花“重瓣化”的花器官形态发育比较观察[J]. 植物分类与资源学报, 2014, 36(3): 303-309.
[11]姜莉, 陈发棣, 滕年, 等. 重瓣和重台莲品种花芽分化过程的解剖结构比较[J]. 园艺学报, 2009, 36 (8): 1233 -1238.
[12]黄国振. 荷花的重瓣化及其遗传基础的初步探讨[J]. 武汉植物学研究, 1983, 1(2): 139-142.
[13]黄国振, 何子灿, 陈纯章, 等. 荷花的重瓣化与染色体组型变异的相关性[J]. 武汉植物学研究, 1983, 1(2): 11-16.
[14]ZHANG W W, TIAN D K, HUANG X, et al. Characterization of flower-bud transcriptome and development of genic SSR markers in Asian lotus (Nelumbo nuciferaGaertn.) [J]. PLoS One, 2014, 7(9):e112223.
[15]COEN E S, MEYEROWITZ E M. The war of the whorls: genetic interactions controlling flower development [J]. Nature, 1991, 353: 31-37.
[16]ANGENENT G C, FRANKEN J, BUSSCHRE M, et al. A novel class of MADS box genes is involved in ovule development in petunia [J].Plant Cell, 1995, 7(10): 1569-1582.
[17]赵印泉, 刘青林. 重瓣花的形成机理及遗传特性研究进展[J]. 西北植物学报, 2009, 29(4): 832-841.
[18]KIM S, KOH J, YOO M J, et al. Expression of floral MADS-box genes in basal angiosperms: Implications on evolution of floral regulators and the perianth [J]. Plant Journal, 2005, 5(43): 724-744.
猜你喜欢
轻音乐(2022年3期)2022-12-29
经济林研究(2022年2期)2022-07-20
课外生活·趣知识(2022年6期)2022-07-14
现代农业科技(2022年4期)2022-03-08
共产党员(辽宁)(2021年17期)2021-10-07
食品与生物技术学报(2021年8期)2021-01-17
中国防伪报道(2018年9期)2018-01-22
现代园艺(2017年1期)2017-07-14
绿色科技(2017年2期)2017-03-23
农村百事通(2016年7期)2016-05-14
