
绵枣儿花部综合特征与繁育系统
2018-07-07 03:47
中南林业科技大学学报 2018年6期
(东北林业大学 园林学院,黑龙江 哈尔滨 150040)
绵枣儿Barnardia japonica Thunb.,也就是以前学者常常称为Scilla scilloides Druce的植物,在较新的分类系统中属于百合科Liliaceae绵枣儿属Barnardia[1],绵枣儿属植物全球约有90种,广泛分布于亚洲、欧洲和非洲的温带地区,少数也分布于热带山地,南非至少具有6种绵枣儿。绵枣儿在我国南北各省均有分布,且在我国只有1个种和1个变种,即绵枣儿和白绿绵枣儿。绵枣儿是一种极具发展、应用潜力的野生植物种类,其花期长,花色艳丽,线条柔美,单株花序平均单花数量100朵以上,是良好的花境、花坛和切花素材。绵枣儿可群植于林间草地、花境和坡地,也可以在山石旁三五株成丛进行点缀,与其他园林景观植物配合,共同营造与众不同的季相景观[2]。且绵枣儿在全国分布范围极广,耐寒、耐旱能力强,生长季节不需要特殊管理,病虫害较少,具有固土的作用,也有一定的生态效益。在园林中最常用的种类为聚铃花和地中海绵枣儿[3]。
自然界仍然存在很多极具观赏价值的植物被人们所忽略,若通过一定研究将它们应用到城市园林建设中,必定会创造出更加多样的园林景观,而绵枣儿就是极具开发潜力的一个,且绵枣儿属下还具有许多观赏价值高的植物种。目前有关绵枣儿属植物的研究,主要进行的是其遗传学[4-5]、育种繁殖[6-7]、细胞学[8-9]、药学[10-11]和化学方面[12-14]的研究,关于绵枣儿属花部综合特征和植物繁育系统方面的相关研究很少,目前国内外只涉及对绵枣儿属开花生物学的初步研究。该研究以从长白山引至东北林业大学园林学院苗圃中的绵枣儿作为研究对象,主要对其花部综合特征、开花动态、繁育系统等方面展开研究,以期为今后绵枣儿的常规育种、杂交育种提供重要的数据支持和理论依据,对绵枣儿属植物新品种的选育提供技术支持,加快绵枣儿在城市园林中的应用。
1 材料与方法
1.1 试验材料
试验材料为从吉林省长白山引种至东北林业大学园林学院苗圃的绵枣儿植株。
1.2 试验方法
1.2.1 绵枣儿花部综合特征的观察
在绵枣儿的盛花期,随机选取30朵正在开放的小花,观察时记录花朵的结构组成、花朵各器官的变化时间及特点,并用游标卡尺来测量其花被片的长和宽、花冠的直径以及雌蕊和雄蕊各部分的长度。
用FAA固定液对现取的花药进行固定和保存,并利用爱氏苏木精染色法进行染色[15],经过脱水、透明、包埋、切片等一系列处理,切片厚度为 8 μm,在LEICA DM 2500显微镜下观察花药横切结构并拍照。
采摘新鲜的绵枣儿花朵,适当清洁表面,分别取花粉、柱头、花瓣、子房进行制样,观察花粉与柱头的结构特征及花瓣与子房的形态,在日立S-520扫描电镜下逐个观察已经经过粘贴喷金等专业处理后的材料,选取不同角度与倍数进行观察拍照。
1.2.2 绵枣儿开花动态观察
(1)单花开放动态:在绵枣儿的花蕾期,随机选取15株植物进行挂牌标记,连续数日采取定位观察并随时记录单花个体的开放过程,直至花朵凋谢。
(2)单株开放动态:在绵枣儿的花蕾期,随机选取50株植物进行挂牌并标明序号,观察并记录单株绵枣儿的花序开放顺序、开花持续的时间以及花部形态特征。始花日期是指单株植物盛开的单花数量达到植株的25%时的日期,盛开的单花数大于或等于50%时即为盛花期,当盛开的单花数小于10%时为末花期,没有花开放时即为终花期。以上数据的平均值作为个体的开花参数。
1.2.3 绵枣儿花粉-胚珠比的检测
花粉-胚珠比的估算采用血球计数器法。随机采取绵枣儿植株上刚刚开放且花药未开裂的小花15朵,选取充分成熟的花药90枚,平均放入3个青霉素小瓶内。待花药自然干燥、开裂并充分散粉时,向每个小瓶内滴进2 mL1%的六偏磷酸钠,盖上盖子并充分震荡,使得花粒处于悬浮状态。吸取1滴悬浮液滴于血球计数板上,在显微镜下观察并记录结果。进行5次重复试验,取结果的平均值。在显微镜下将单花子房进行解剖,并统计胚珠的数量。采用Cruden[16]方法进行花粉-胚珠比的估算(P/O),即P/O=单花花粉量/胚珠数。
1.2.4 绵枣儿杂交指数的估算
对绵枣儿的单花直径进行测量,并观察绵枣儿的开花行为,运用Dafni[17]的标准来判断绵枣儿的繁育系统。判断方法为:(1)单花直径>6 mm则观测值为3;2~6 mm则观测值为2;1~2 mm则观测值为1;<1 mm则观测值为0。(2)花药开裂期与柱头可授期之间是否存在时间上的间隔,雄蕊先开裂则观测值为1;雌蕊先具有可授性或者雌雄蕊同时成熟则观测值为0。(3)花药与柱头之间的空间关系,二者空间分离则观测值为1;二者同一水平则观测值为0。这3个指标的和即为绵枣儿的杂交指数(OCI)。
评判绵枣儿繁育系统类型的标准为:OCI和为4,则该植物的繁育系统为部分可以自交亲和,异交为主,需要传粉者进行传粉;OCI和为3时,该植物的繁育系统为自交亲和,少数情况需要传粉者的参与;OCI和为2,该植物的繁育系统为兼性自交型;OCI和为1时,该繁育系统为专性自交型;OCI和为0时,则被称作闭花受精型的繁育系统。
1.2.5 绵枣儿套袋、去雄以及人工授粉试验
随机选取即将开放的单花进行人工授粉试验,共8种处理,每个处理标记50朵单花,按照Dafni[17]记载的试验方法对单花进行以下处理:(1)自然授粉,不去雄,不套袋。(2)自然授粉,不去雄,花朵开放前使用细白纱网套袋。(3)自然授粉,不去雄,套袋。(4)自然授粉,去雄,不套袋。(5)人工自花授粉,去雄,套袋。(6)人工进行同株异花授粉,并去雄、套袋。(7)人工进行异株授粉,去雄,套袋。(8)不授粉,去雄,套袋。
1.2.6 数据分析
运用SPSS 19.0软件进行数据统计与分析,用Excel软件进行数据初步处理并绘图。
2 结果与分析
2.1 花部综合特征观察
2.1.1 绵枣儿花部形态及结构
绵枣儿为多年生草本植物,株高20~40 cm,具有卵球形的鳞茎。多数具有叶片3~6枚,叶狭线形,长15~30 cm,深绿色,革质。花葶30~45 cm,直立,高出叶丛,顶生总状花序,长24~38 cm,每个花序具有小花40~300朵不等,线状苞片长1~2 mm。花被片6枚,且略向外翻卷,具有1条绿色脉纹,花色为淡紫红色、淡粉色。绵枣儿的花为两性花,丁字着药,每朵花具有6枚花药(图1,A),花药四室(图1,B),左右对称分布,中央通过药隔相互连接,花药横切面近似四边形或蝴蝶形(图1,C)。
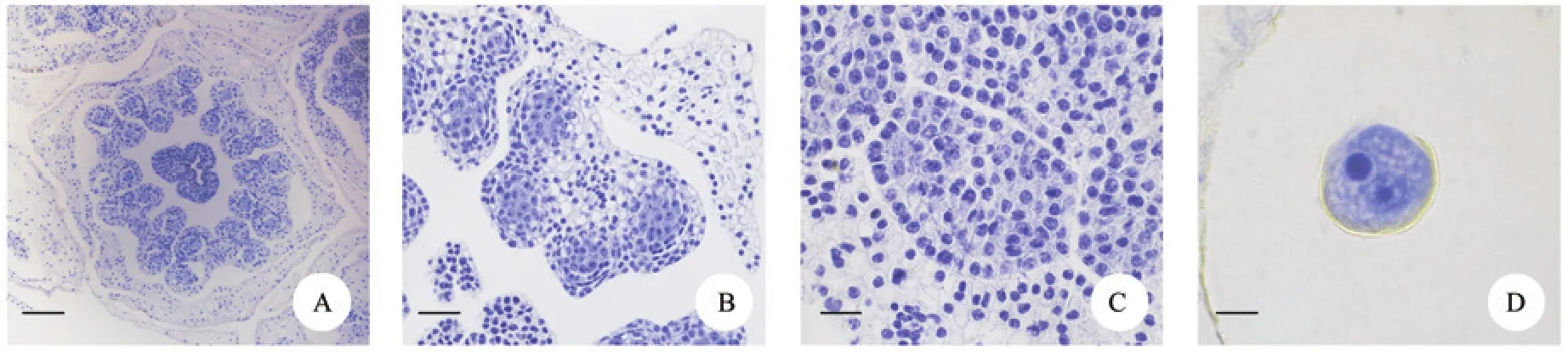
图1 绵枣儿单花花药横切面结构Fig.1 Cross section view of anther of Barnardia japonica
绵枣儿花朵开放时(表1),花朵淡紫红色,直径为7.12~9.02 mm;花被片长3.54~4.52 mm,宽1.20~1.96 mm;花丝白色,基部扁平,先端尖,长2.76~3.16 mm;花药初期颜色为黄绿色,后逐渐变成灰绿色,长1.14~1.46 mm,宽0.70~1.00 mm;花柱高0.58~1.04 mm,乳白色,柱头表面呈乳突状,成熟时分泌少量粘液;子房为椭圆形,长1.98~2.38 mm,宽0.58~1.78 mm,淡粉色,授粉后颜色加深,最终为绿色。花瓣基部具有分泌物(表2)。

表1 绵枣儿花部器官数量特征统计值Table 1 Statistics on the number of flower organs of Barnardia japonica

表2 绵枣儿花朵形态特征Table 2 The floral morphology of Barnardia japonica
2.1.2 绵枣儿花部微观形态观察
当花药开裂后,花粉散出,成熟绵枣儿花粉内只含有营养细胞和生殖细胞,生殖细胞没有进行有丝分裂而产生精细胞,因此,绵枣儿被划分为二细胞型(图1,D)。花粉为单粒,超长球形(图2,A)。极面观为近圆形(图2,D),赤道面观为舟形(图2,C)。花粉粒大小约为48.75 μm×20.48 μm。根据Erdtman的标准进行判断,绵枣儿的花粉为中等孢粉。花粉的萌发器官是单沟,其萌发沟宽而浅,延长至两端(图2,B)。花粉外壁光滑,具有不规则的小孔,小孔分布较均匀(图2,E)。花粉萌发时,大多数花粉能萌发出1个花粉管,部分花粉可萌发出2个花粉管(图2,P-Q)。绵枣儿柱头的相对表面积较大,表面为许多球状凸起(图2,F-G)。柱头成熟时有少量粘液出现,柱头伸长,可以粘附和嵌入花粉(图2,H-I,S-T)。绵枣儿花粉较大,不具备气囊,同时柱头基本与花药高度持平。而风媒植物花粉质轻、干燥,通常具有气囊,柱头一般无阻碍的暴露在外面,雌蕊较长。根据上述特点,绵枣儿的花粉和柱头的形态特征与虫媒花的特点相符合,属于虫媒植物。
绵枣儿的子房呈椭球形,基部略宽,呈淡粉红色,有6条纵白色棱(图2,R)。通过扫描电镜对子房外表面进行观察,可见其表面呈鳞片状,均匀致密,鳞片表面布满不规则的片状凸起(图2,J-K)。

绵枣儿花被片为淡紫红色,共6枚,花被片表面光滑,由不规则的细胞构成,细胞表面具有条状凸起(图2,L)。花被片上可见随机分布的气孔,气孔长约19.8 μm(图2,M-O)。

图2 绵枣儿花粉和柱头的微观形态Fig.2 Microstructure of dust and stigma of Barnardia japonica
2.2 绵枣儿开花动态
2.2.1 单花开放动态
绵枣儿的花蕾为绿色,逐渐膨大,到开花前顶端变为淡紫红色,此时花蕾直径达到2.49 mm左右。花朵开放过程中,花蕾顶端松动,一侧的3枚花被片先展开,另外3枚随后展开,至6枚花被片完全伸展需要0.5~1 h。花朵刚开放时柱头略低于花药或与花药高度持平。开花1~2 d ,粘液量分泌达到最大。花丝在开放过程中,柱头伸长,且张开角度增加,花药在花被片完全打开后逐渐开裂,无先后次序,花药暴露于空气中20~40 min内便开始开裂,6枚花药完全开裂需要1~2 h,当天基本可以完成散粉。
绵枣儿花朵开放集中于每天的10:00—15:00之间,夜间不开花,且夜间花被片不闭合,阴雨天也可以开花。在开花前1~2 h花蕾开始松动,当日即可完成开花,柱头分泌粘液,花药完全开裂。至第4或5 d,花瓣皱缩,雄蕊、雌蕊皆开始变形枯萎,花朵开始萎蔫。
绵枣儿的单花持续时间为4~5 d,其单花期根据花部形态变化和散粉特点可以分为3个时期:(1)散粉期:花蕾松动、花被片开展至6个花药完全开裂,柱头无明显变化;(2)柱头发育期:柱头伸长,颜色由白色变为乳白色转黄,最终变为黄褐色,且柱头弯曲,顶端具有乳突。(3)凋谢期:开花4~5 d,花瓣开始皱缩,花丝萎蔫,花药脱落,柱头枯黄(图3)。

图3 绵枣儿单花开放动态Fig.3 Open dynamic of single fl ower of Barnardia japonica
2.2.2 单株开放动态
单株开放具有一定的次序,为由下至上逐渐开放,绵枣儿经历了花蕾发育、始花期、盛花期、末花期、果实发育期和种子成熟期(图4)。一个花序从冒出地面开始直至开出第一朵小花需要经过10~20 d,每个花序具有40~300个花蕾不等。通过对两年植株开花动态的观察,发现绵枣儿单株花期一般在9~28 d之间波动,这一般取决于每株小花数量的多少。在2015年的单株花期中,16 d的分布频率最高,达到20%,17 d次之,达到总频率的14%,而在2016年的单花花期中,14 d的分布频率最高,为22%,16 d次之,达到14%(图5)。开花的过程中,随着时间推移,花朵开放百分率呈现出单峰式变化规律,单株高峰出现在5~9 d之间,花朵开放百分率最高可达8.9%。开花式样也同样呈现出明显的单峰式变化趋势,单株花期的前期和后期大部分时期,开花样式很小,开花式样最大的一天出现在开花8~11 d之间,统计50个单株的开花式样,平均最大量为60朵,占整个花序的49.2%。

图4 绵枣儿单株开放动态Fig.4 Development of individual opening of Barnardia japonica
2.3 绵枣儿繁育系统的确定
2.3.1 花粉-胚珠比
绵枣儿花的单花花粉数平均为9 333粒,单花胚珠数为3个,P/O约为3 111,根据Cruden[16]关于繁育系统的判断标准,绵枣儿的繁育系统属于专性异交类型。

图5 绵枣儿花序开放持续时间的频率分布Fig.5 The frequency distribution of inf l orescences opening duration of Barnardia japonica
2.3.2 杂交指数
根据Dafni[17]的方法估算绵枣儿的杂交指数。结果为:绵枣儿花朵直径8 mm,大于6 mm记为3;雌蕊在花药初期进行散粉时已经具备了一定的可授性,可以认为雌蕊和雄蕊在成熟的时间上同步,记为0;花柱与花药在花开后空间上始终分离,记为1。综合以上结果OCI值等于4,即绵枣儿的繁育系统主要以异交为主,部分可以自交亲和,需要传粉者的参与。
2.3.3 套袋试验
通过对绵枣儿进行套袋授粉试验,检测绵枣儿的授粉方式(表3)。自然状态下绵枣儿的坐果率为44.0%,结籽率为20.6%,经过套袋试验发现,人工授异株花粉的坐果率可高达78.0%,而其他方式较低,结合上述结果可断定绵枣儿的繁育体系以异交为主。

表3 绵枣儿套袋试验结果Table 3 The test results of emasculation of Barnardia japonica
按照Dafni[17]的方式测量可以判断绵枣儿的繁育系统以异交为主,部分可以自交亲和,需要传粉者的参与;依据Cruden[16]对植物花粉-胚珠比的划分标准,绵枣儿繁育系统为异交类型;通过套袋试验得出,绵枣儿的繁育体系以异交为主。综合以上结果推断,绵枣儿属于以异花传粉为主的植物,部分自交亲和,传粉过程中需要传粉者的参与。
3 结论与讨论
植物的营养状况、传粉机制、传粉行为以及对外界环境条件的适应度都会影响花部的综合特征[18-19]。绵枣儿的种群花期为7月中下旬至8月中下旬,花期较短,为了能够在炎热的夏季达到生殖成功的目的,绵枣儿花展开了一系列吸引传粉者的策略。绵枣儿花为顶生总状花序,长24~38 cm,每个花序均具有小花40~300朵不等,且由下至上逐渐开放,绵枣儿的单花开放持续时间为4~5 d,单株花期可以达到9~28 d。绵枣儿开花量较大且单花开放不同步的这个特性对于增加昆虫的访花行为、保证其有足够的时间完成传粉受精具有积极意义。绵枣儿的花色为淡紫红色、淡粉色,花色明显,总状花序上的每朵小花都具有花药6枚,绵枣儿明亮的花色以及较多的花粉和花蜜可以吸引更多的昆虫传粉,从而增加植株的授粉结实机会。柱头上的乳突与分泌物有助于花粉的萌发,绵枣儿的柱头表面有许多球状凸起,相对表面积较大,柱头成熟时有少量粘液出现,柱头伸长,这有利于花粉萌发与昆虫进行传粉活动[20]。
在自然界中,因为异交有利于提高植物的遗传多样性,植物通常会通过雌雄异熟、雌雄异位、自交不亲和机制等多种方式来保证植物异株异花授粉,以确保其后代能够更好地来适应环境[21]。绵枣儿的花为两性花,丁字着药,绵枣儿6枚花药完全开裂后花柱才开始伸长,其花药与柱头在开花前后空间上始终分离,说明绵枣儿单花自交受到一定程度的限制,需要传粉者。绵枣儿的花粉粒大小约为48.75 μm×20.48 μm,属于中等孢粉,并且不具备气囊,不易被风吹走,在自然条件下,须有传粉者才能完成授粉过程。绵枣儿的这些花部特征均有利于其异花授粉成功,这也是绵枣儿长期繁殖进化的一种成功的适应机制。
P/O值是衡量植物对雄性资源分配量的一个标准,当植物的P/O值越高时,其传粉的效率就越低,基本的繁育系统类型可以被预知[22]。本研究表明绵枣儿单花花粉量较大,但胚珠只有3个,P/O值约为3 111,P/O值较高,较大程度上限制了绵枣儿的自花传粉,需要通过异交来提高它的授粉率。根据Dafni[17]的方法对绵枣儿的杂交指数进行测量,绵枣儿的OCI值为4,进一步证明了绵枣儿的繁育系统类型是以异交为主,部分植株自交也可以亲和,但是需要传粉者的参与。
在外界的自然条件下,植物的自交率受到多种因素的干涉,植物本身和其他个体的花粉数量与质量、植物在进行受精时异花花粉对自花花粉形成的竞争强弱以及其他个体外来花粉的授粉时间都会影响植物的自交率[23-26]。自然状态下绵枣儿的坐果率为44.0%,结籽率为20.6%,均较低,这说明绵枣儿花存在一定的自花限制,造成这一现象的主要原因可能是由于其花药与柱头在开花前后空间上始终分离,花粉与柱头缺乏有效触碰。在去雄条件下对绵枣儿进行自由授粉,其花座果率是48%,结籽率是27.8%,这说明在自然条件下植物传粉媒介的效率能够影响植物异花授粉的结果。绵枣儿在人工同株异花授粉的情况下,坐果率仅仅为32%,这表明绵枣儿自交亲和率较低。人工授粉以及套袋试验发现,人工授异株花粉的座果率可高达78.0%,与其他方式相比较高,由此可断定绵枣儿的繁育体系以异交为主,需要传粉者,再一次验证上述绵枣儿的繁育系统以异交为主的判断。
植物的开花机制不仅受温度、光照、水分、土壤等环境因素的影响,还会受到植物自身生理学机制及分子学机制等遗传条件的制约[27-28]。本试验对绵枣儿的花部综合特征与繁育系统进行了较为细致的研究,对认识绵枣儿的开花生物学特性和整个开花过程的自然规律具有一定积极影响。目前,国内外开展绵枣儿遗传基因对其开花机制影响的研究甚少,在后期研究中将对绵枣儿开花的分子生物学机制进行大量研究,进一步从分子水平上研究绵枣儿的开花机制,以期用分子技术加快绵枣儿属植物新品种的选育和绵枣儿在城市园林景观中的应用[29]。
[1]Angiosperm Phylogeny Group.An update of the Angiosperm Phylogeny Group classification for the orders and families of flowering plants: APG IV[J].Botanical Journal of the Linnean Society,2016,181 (1): 1-20.
[2]姚悦梅,潘耀平,毛忠良.绵枣儿入园林[J].植物学杂志,2002(5):23.
[3]Crouch N R, Bangani V, Mulholland D A.Homoisoflavanones from three South African Scilla species[J].Phytochemistry,1999,51(7):94-946.
[4]丁开宇,王 伟,孙静贤,等.Giemsa-C带揭示的绵枣儿多倍体复合体染色体变异[J].武汉植物学研究,2007,25(5):421-426.
[5]尚彩玲,丁开宇.绵枣儿核核糖体DNAITS序列扩增的研究[J].山西中医学院学报,2009,10(4):52-54.
[6]顾福根,万志刚,赵旭平.地中海绵枣儿的组织培养与植株再生[J].植物生理学通讯,2007,43(1):118.
[7]翟素萍,熊 丽,王继华,等.绵枣儿叶片植株再生体系的建立[J].植物生理学通讯,2006,42(1):69.
[8]Araki H.The distribution of diploids and polyploids of the Scilla scilloides complex in Korea[J].Genetica,1985,66(1):3-10.
[9]Yü Zhi-hui, Araki H.The distribution of diploids and polyploids of the Scilla scilloides complex in the northeastern district of China[J].Journal of Plant Research,1991,104(3):183-190.
[10]Dias C, Graca J A B, Goncalves M L.Scilla maderensis, TLC screening and positive inotropic effect of bulb extracts[J].Journal of ethnopharmacology,2000,71(3):487-492.
[11]Sparg S G, Staden J Van, Jäger A K.Pharmacological and phytochemical screening of two Hyacinthaceae species:Scilla natalensis and Ledebouria ovatifolia[J].Journal of Ethnopharmacology, 2002,80(1):95-101.
[12]Asano N, Ikeda K, Kasahara M, et al.Glycosidase-Inhibiting Pyrrolidines and Pyrrolizidines with a Long Side Chain in Scilla peruviana[J].Journal of natural products,2004,67(5):846-850.
[13]Kato A, Kato N, Adachi I, et al.Isolation of glycosidaseinhibiting hyacinthacines and related alkaloids from Scilla socialis[J].Journal of natural products,2007,70(6):993-997.
[14]任福才,王丽霞,王 飞,等.绵枣儿化学成分研究[J].中草药, 2014,45(14):1984-1988.
[15]张志良,瞿伟菁.植物生理学实验指导[M].北京:高等教育出版社,2003,127-128.
[16]Cruden R W.Pollen-ovule ratios: A conservative indicator of breeding systems in flowering plants[J].Evolution, 1997,31(1):32-46.
[17]Dafni A.Pollination ecology: A practical approach[J].New York:Oxford University Press, 1992,1-57.
[18]Wyatt R.Pollinator-plant interactions and the evolution of breeding systems[J].Pollination biology,1983,51:96.
[19]杨 亚,袁德义,刘东明,等.不同授粉方式对攸县油茶坐果率及果实生长的影响[J].中南林业科技大学学报,2016, 36(2):61-65.
[20]徐丽萍,喻方圆.东京野茉莉雌雄蕊的形态及显微结构[J].南京林业大学学报( 自然科学版), 2017,41(2):34-40.
[21]刘 鹏,刘惠民,王连春,等.牛角瓜花部综合特征与繁育系统[J].园艺学报, 2015,42(10):2002-2014.
[22]李志成,李 进,吕海英,等.艾比湖沙拐枣的花部特征与繁育系统[J].园艺学报,2015, 42(5):939-949.
[23]Primack R B.Longevity of individual fl owers[J].Annual Review of Ecology and Systematics, 1985, 16:15-37.
[24]Schoen D J, Ashman T L.The evolution of flower longevity:Resource allocation to maintenance versus construction of repeated parts in modular organisms[J].Evolution, 1995, 49:131-139.
[25]朱江华,吕芳德,王 森,等.6个蓝莓品种花粉特性和花粉育性的比较[J].经济林研究,2016,34(2):101-108.
[26]苏 瑾,芦 娟,吴文俊,等.6个油橄榄品种的花粉特性[J].经济林研究,2017,35(3):208-212.
[27]冯贝贝,魏雅君,李雯雯,等.阿克苏地区3个欧洲李品种花芽分化的进程[J].经济林研究,2017,35(3):156-160.
[28]王丽媛,郭素娟.板栗‘燕山早丰’雌花分化的解剖学研究[J].中南林业科技大学学报,2017,37(1):43-47.
[29]宋 猜,姜仲茂,李 慧,等.仁用杏花芽3个发育时期转录组表达谱研究[J].经济林研究,2016,34(4):14-19.
猜你喜欢
安徽农业科学(2022年18期)2022-10-13
麦类作物学报(2022年8期)2022-08-08
新疆农垦科技(2022年1期)2022-03-10
华东科技(2021年9期)2021-09-23
作文成功之路·中考冲刺(2021年3期)2021-05-07
学生天地·初中(2019年8期)2019-10-08
学生天地(2019年22期)2019-09-26
蔬菜(2019年5期)2019-05-21
小雪花·小学生快乐作文(2018年9期)2018-02-19
江苏农业科学(2016年8期)2017-02-15
