
不同放牧制度对高原鼢鼠活动区高寒草地群落结构影响
2018-07-30 03:28王玉琴鲍根生王宏生李继鹏
草地学报 2018年1期
王玉琴, 鲍根生, 王宏生, 曾 辉, 李继鹏
(1. 省部共建三江源生态与高原农牧业国家重点实验室,青海大学畜牧兽医科学院,青海 西宁810016; 2. 青海省青藏高原优良牧草种质资源利用重点实验室,青海省畜牧兽医科学院, 青海 西宁810016)
青藏高原是我国重要的畜牧业基地之一,其中天然草地面积高达1.5亿hm2,是当地畜牧业的发展基本物质保障。同时,青藏高原涵盖多种草地类型,其中高寒草甸是该区域分布面积最广,也是生态功能稳定性较弱的草地生态系统[1-2]。多年来,由于全球气候变化、过度放牧、人类不合理的活动及啮齿动物挖掘活动等因素的综合作用,青藏高原地区约50.4%高寒草地处于退化状态,16.5%高寒草地处于严重退化状态[3-4]。其中,草地载畜量过高是造成高寒草甸退化的主要原因,而大面积出现的退化草地往往成为草原啮齿动物最佳的栖息地[5-6]。高寒草甸主要原生植被-莎草科植物依靠发达的根茎繁殖能力形成紧实致密的草毡层,莎草科植物地上部分被草食家畜反复啃食及啮齿动物大量取食地下根系后,莎草科植被大面积减少,造成草地植被结构快速发生变化、物种多样性快速下降,加之冻融、风雨侵蚀等物理过程,最终形成毒杂草大面积滋生的“黑土滩”退化景观[7-8]。因此,草食家畜采食和啮齿动物挖掘行为对草地的扰动强度在一定程度上决定高寒草地的退化进程和植物群落的演替方向。
啮齿动物是草地生态系统重要的组成部分,通过采食和挖掘活动影响草地植被的分布和组成,进而影响整个草地生态系统[9]。高原鼢鼠(Myospalaxbaileyi)和高原鼠兔(Ochotonacurzoniae)是青藏高原地区主要的啮齿动物种类,高原鼢鼠是一种视力退化、终年营地下生活的啮齿动物,主要以植物根系为食物来源[10],其挖掘和造丘活动将地下新鲜土壤上翻到草地表面,形成大小不一的土丘[11-12],进而影响地上植被空间结构、土壤形态及土壤矿质化进程[13]。青海省高原鼢鼠发生面积为2 271万hm2,危害面积为1 561万hm2[14]。高原鼢鼠大面积发生与草地生态系统被长期扰动有关(如过度放牧、垦荒等),当草地载畜量较低或适中时,高原鼢鼠活动能提高草地群落物种多样性和增强草地生态系统的稳定性,成为“草地生态系统工程师”;然而,草地载畜量持续过高时,高原鼢鼠活动能加速草地群落的逆向演替,导致草地生态系统退化甚至崩溃,间接成为是草地退化的主要驱动力[15]。
合理的放牧制度能提高草地群落物种多样性和草地生态系统的稳定性。关于受损草地生态系统的恢复,最常用的方法有围栏封育(自然恢复),它是人类有意识调节草地生态系统中草食动物与植物的关系以及管理草地的手段[16],具有促进草地生态系统良性循环的功效[17]。单贵莲等[18]研究表明,围栏封育能短期内恢复草地建群种的盖度,而轻度放牧和中度放牧能提高草地群落物种多样性。物种多样性一定程度上能决定高原鼢鼠的食物数量和品质[19-20],也是决定高原鼢鼠选择适合栖息地主要因素。目前,有关不同放牧制度对草地生物多样性的影响[21-22]及高原鼢鼠活动对草地群落演替等方面的相关研究比较详尽[23-25];然而针对高原鼢鼠活动对不同放牧制度下草地群落结构影响的研究尚少有报道。基于此,本研究通过分析不同放牧制度对草地群落多样性、高原鼢鼠相对种群数量的影响及草地群落结构与高原鼢鼠种群数量的相关性,明确不同放牧制度对高原鼢鼠种群数量的调控作用,为青藏高原地区高原鼢鼠的生物防控提供科学依据。
1 材料与方法
1.1 试验区概况
试验点位于青海省黄南藏族自治州河南县托叶玛乡文群村(N 34°37′56″,E 106°23′20.2″),海拔3 480 m,属于典型的高原大陆性气候,具有日照时间长、辐射强、热量低、气温日差较大,冬季寒冷漫长,夏季凉爽短促,冬春季节多霜冻、雪灾、风灾等特点。年均温-1.3℃~1.6℃,年日照时数2 530~3 100 h,年降水量597.33~615.6 mm,年蒸发量为1 349.7 mm。植被类型主要以高寒嵩草草甸为主,土壤类型为典型的高寒草甸土。草地植被的优势种为矮嵩草(Kobresiahumilis)、垂穗披碱草(Elymusnutans)、线叶嵩草(Kobresiacapillifolia),伴生种主要以珠芽蓼(Polygonumviviparum)、密花香薷(Elsholtziadensa)、鹅绒委陵菜(Potentillaanserina)、莓叶萎陵菜(Potentillafragarioides)、蒲公英(Taraxacummongolicum)等杂类草为主。
1.2 样地设置
自2014年在黄南州河南县托叶玛乡文群村高原鼢鼠危害严重的天然草场上,采用网围栏分别建立3个禁牧区和3个季节放牧区,面积为200 m×200 m。季节放牧区主要以当地冬季草场为主,每年11月至第二年5月进行放牧;同时选取水源地周围、全年家畜采食的草地划定为自由放牧区。通过设定禁牧区、季节性放牧区和自由放牧区来模拟家畜对草地的不同利用方式。自由放牧区和季节性放牧区的放牧强度均为4只羊·hm-2。
1.3 测定内容及方法
1.3.1草地群落结构调查 2014年、2015年8月上旬,在不同处理样地内分别随机选取5个样方(0.5 m×0.5 m),统计样方内出现的植物种类,测定植物高度、盖度、地上生物量和地下生物量。其中,采用目测法估测每种植物的盖度;采用分种收获法测定样方内出现各物种的干重[26-27];利用铁锹挖取体积5个体积为20 cm×20 cm×30 cm的土块,用清水清洗去除泥土,依据地上植物形态特征区分各物种并归类,带回试验室70℃烘干称重。
1.3.2草地植物多样性指数的测定 物种重要值(Pi)=(相对高度+相对盖度+相对生物量)/3;
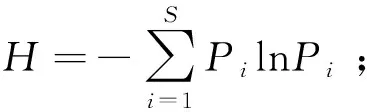

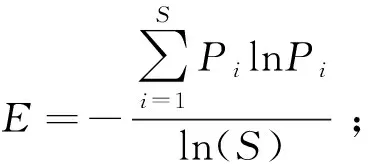
物种丰富度指数(S)=物种数;
式中,Pi=Ni/N,Pi代表物种i的相对重要值,N代表样方各物种重要值总和,Ni代表样方内第i物种的重要值[27]。
1.3.3高原鼢鼠新鼠丘数调查 依据王权业等[14]、刘丽等[28]利用高原鼢鼠在挖掘活动较频繁的春秋两季所形成的新鼠丘数量能可靠估测该区域高原鼢鼠数量的方法,于2014年5月初分别在禁牧区、季节性放牧和自由放牧区选取5个25 m×25 m样区,每个样区间隔500 m,调查不同样区内出现的新鼠丘的数量;其中,判定高原鼢鼠新鼠丘的标准为鼠丘土壤新鲜、鼠丘上无植物定植且鼠丘表面出现龟裂。
1.4 数据分析
禁牧区、季节性放牧区和自由放牧区内出现的鼢鼠新鼠丘数量、生物量(地下、地上)及植物群落生物多样性指数采用单因素方差分析,分别比较不同样区内各测定指标间的差异;其中,植物群落多样性指数采用独立样本T检验分别检测2014、2015年间同一样区间的差异。同时,采用线性回归分析的方法,分析不同样区内鼢鼠新鼠丘数量与单子叶、双子叶及总生物量的相关性,采用SPSS 17.0软件进行分析。
2 结果与分析
2.1 高原鼢鼠活动对不同放牧制度样地物种重要值的影响
高原鼢鼠活动对不同放牧制度的草地群落各物种的组成和重要值变化有一定的影响(表1)。总体表现为:季节性放牧区两年间物种数量平均值(36种)高于禁牧区(30种)和自由放牧区(32种),同时季节性放牧区草地建群种[禾本科(Grmineae)、莎草科(Cyperaceae)]的重要值高于自由放牧区和禁牧区;而阔叶类植物在不同放牧制度样地间存在差异,如禁牧区内玄参科(Scrophuariaceae)和菊科(Asteraceae)植物的重要值较高,蔷薇科(Rosaceae)植物在自由放牧区内的重要值较高,而季节放牧区内龙胆科(Gentianaceae)植物的重要值较高。
由表1可知,2014-2015年间不同放牧制度样地内草地群落中垂穗披碱草的重要值最高,且禁牧区内垂穗披碱草的重要值明显高于自由放牧区和自由放牧区;禁牧两年后草地群落中矮嵩草、早熟禾、中国马先蒿、肉果草、冷蒿、老鹳草、珠芽蓼、密花香薷和田葛缕子的重要值增加,而垂穗披碱草、鹅绒委陵菜、莓叶委陵菜、美丽风毛菊、车前、多枝黄芪和蒲公英的重要值降低;自由放牧方式下草地群落中矮嵩草、早熟禾、碎米蕨叶马先蒿、鹅绒委陵菜、莓叶委陵菜、黄帚橐吾、冷蒿、毛茛、密花香薷和獐牙菜的重要值比2014年增加,而垂穗披碱草、芒冾草、中国马先蒿、肉果草、美丽风毛菊、车前、多枝黄芪和蒲公英的重要值比2014年降低。季节放牧区中早熟禾、米口袋、金莲花、扁蕾、龙胆、碎米蕨叶马先蒿、莓叶委陵菜、肉果草、多枝黄芪、冷蒿、老鹳草和毛茛的重要值较2014年有所增加,而垂穗披碱草、矮嵩草、线叶嵩草、獐牙菜、芒冾草、鹅绒委陵菜、珠芽蓼和密花香薷的重要值较2014年有所降低。

表1 不同放牧制度的草地群落主要物种重要值Table 1 Species importance values of grassland community in different grazing systems
2.2 高原鼢鼠活动对不同放牧制度的草地群落多样性影响
高原鼢鼠活动对不同放牧制度下草地群落的香农威纳指数、优势度指数、均匀度指数和物种丰富度指数产生不同程度的影响(表2)。从年度变化来看,禁牧、季节性放牧和自由放牧区的香农威纳指数、优势度指数和物种丰富度指数在第二年增加;同时,季节性放牧区内第二年草地群落的均匀度指数显著高于第一年(P<0.05)。从不同放牧制度来看,自由放牧区第一年的优势度指数和均匀度指数高于季节放牧区和禁牧区;而季节性放牧第二年草地群落多样性指数高于禁牧区和自由放牧区,同时季节性放牧区的优势度指数显著高于禁牧区和自由放牧区(P<0.05)。

表2 不同放牧方式下草地群落多样性指数Table 2 Diversity indexes of grassland community in different grazing systems
注:不同字母表示不同放牧制度间差异显著(P<0.05);*表示不同年份间差异显著(P<0.05)
Notes:Different lowercase letters indicate significant differences among different grazing systems (P<0.05);*indicates significant differences among different years at the 0.05 level
2.3 高原鼢鼠活动对不同放牧制度的草地群落生物量的影响
高原鼢鼠活动对不同放牧制度下草地群落地上、地下生物量分配产生了一定程度的影响(图1)。其中,地上生物量表现为:禁牧区>季节性放牧区>自由放牧;而地下生物量表现为:自由放牧区>禁牧区>季节性放牧区。按单子叶和阔叶类植物区分,单子叶植物地上生物量表现为:禁牧区>自由放牧区>季节性放牧区,且禁牧区显著高于季节性放牧区和自由放牧区(P<0.05);地下生物量表现为:自由放牧区>禁牧区>季节性放牧区。季节性放牧区阔叶类植物地上和地下生物量均高于禁牧区和自由放牧区,且禁牧区的地下生物量显著低于季节放牧区和自由放牧区(P<0.05)。

图1 不同放牧制度对草地群落生物量的影响Fig.1 Effects of different grazing systems on grassland community biomass 注:不同小写字母表示不同放牧制度之间草地地上生物量差异显著;不同大写字母表示不同放牧制度之间群落地下生物量差异显著 Note:Different lowercase letters indicate significant difference of above-ground biomass among different grazing systems at the 0.05 level;different capital letters indicate significant difference of below-ground biomass among different grazing systems at the 0.05 level
2.4 高原鼢鼠相对鼠群密度与植物地上、地下生物量的相关性
不同放牧制度下高原鼢鼠相对种群密度存在差异,其中,季节放牧区和自由放牧区的鼢鼠相对种群密度显著高于禁牧区(P<0.05,图2)。同时,高原鼢鼠相对鼠群密度与不同放牧制度样地植物的地上、地下生物量存在相关性(表3)。禁牧区高原鼢鼠相对种群密度与单子叶植物地上生物量、阔叶类植物地下生物量呈正相关,而与单子叶植物地下生物呈显著的负相关(表3);自由放牧区的相对鼠群密度与单子叶植物地上生物量呈负相关,而与单子叶植物地下生物量、阔叶类植物生物量(地下、地上)均呈正相关;季节性放牧区的相对鼠群密度与单子叶、阔叶类植物地上生物量呈负相关,而与地下生物量呈正相关。

图2 不同放牧制度样地相对高原鼢鼠密度Fig.2 Relative density of plateau zokor in different grazing systems
表3 不同放牧制度样地高原鼢鼠鼠群密度与生物量的相关性Table 3 The correlation between relative density of Myospalai baileyi and biomass in different grazing systems
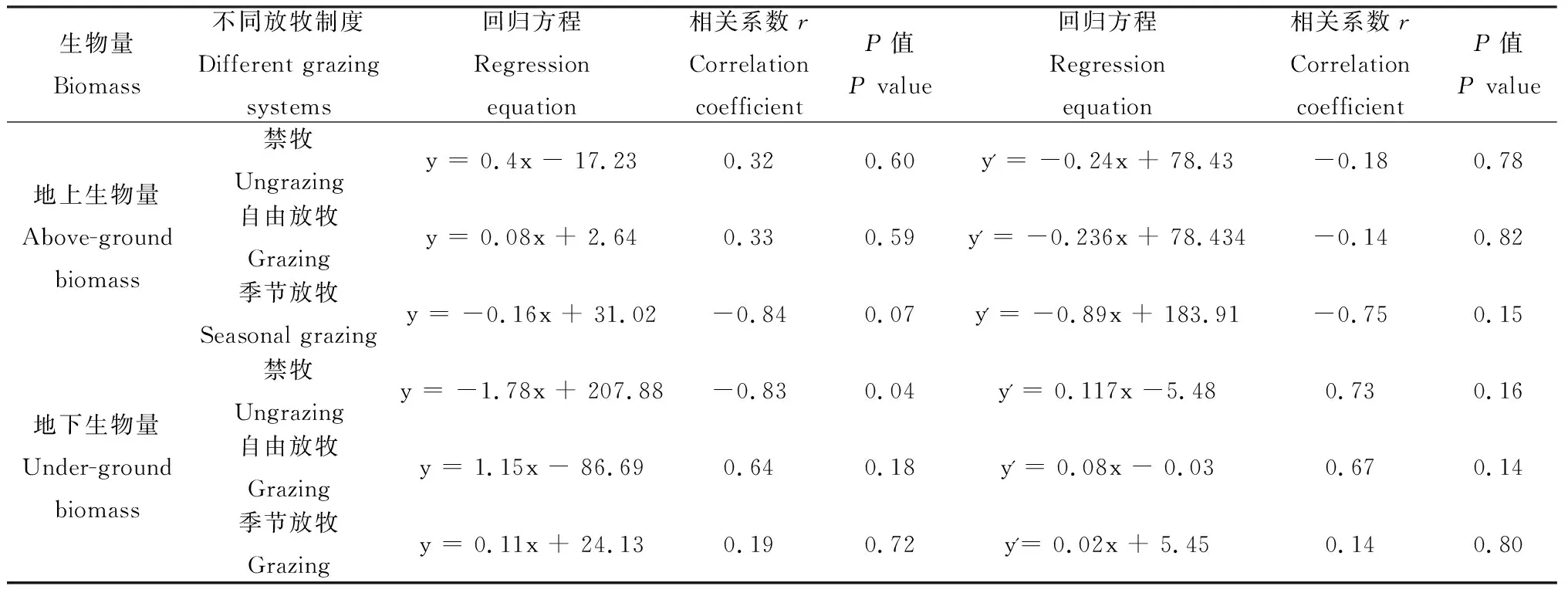
生物量Biomass不同放牧制度Different grazing systems回归方程Regression equation相关系数rCorrelation coefficientP值P value回归方程Regression equation相关系数rCorrelation coefficientP值P value地上生物量Above-groundbiomass禁牧Ungrazingy = 0.4x - 17.230.320.60y' = -0.24x + 78.43-0.180.78自由放牧Grazingy = 0.08x + 2.640.330.59y' = -0.236x + 78.434-0.140.82季节放牧Seasonal grazingy = -0.16x + 31.02-0.840.07y' = -0.89x + 183.91-0.750.15地下生物量Under-ground biomass禁牧Ungrazingy = -1.78x + 207.88-0.830.04y' = 0.117x -5.480.730.16自由放牧Grazingy = 1.15x - 86.690.640.18y' = 0.08x - 0.030.670.14季节放牧Grazingy = 0.11x + 24.130.190.72y'= 0.02x + 5.450.140.80
注:y 表示单子叶生物量;y'表示阔叶类生物量;x表示鼢鼠相对鼠群密度
Note: y indicates biomass of monocotyledons; y' indicates biomass of dicotyledons; x indicates the relative density ofMyospalaibaileyi
3 讨论与结论
草食家畜和啮齿动物作为草地主要的植食性动物,其采食、挖掘、践踏等行为影响草地生态系统结构和功能的稳定性,而草食动物种群数量和分布也受到草地初级生产力的严格限制[29]。由于青藏高原地理环境的特殊性和气候的复杂性,造成青藏高原高寒草甸生态系统具有不稳定性和脆弱性。近年来,由于生物和非生物因素的综合作用,造成高寒草甸生态系统被破坏甚至崩溃,而草地载畜量过高和啮齿动物的破坏是造成这一现象的主要原因[15,30]。家畜和啮齿动物通过地上、地下同时采食,造成植物生长更新进程抑制或停止,进而影响植物群落组成、草地群落逆向演替[31]。通过建立合理的放牧制度一方面能确保草地生态系统的稳定性,另一方面能提高草地的利用率[17]。买小虎等[32]和彭祺等[33]人研究表明,季节性放牧能优化草地群落结构,提高物种多样性。樊江立等[34]研究表明,中度放牧干扰能提高草地群落多样性,而持续高强度放牧干扰后草地群落多样性降低。本研究也得到相同的结论,季节性放牧区物种数、草地建群种的重要值、草地群落多样性指数以及优势度指数均高于禁牧区和自由放牧区,而自由放牧区保持较低多样性,该结果也与Connell提出的“中度干扰理论”假说一致,中等程度的干扰水平能保持较高物种多样性和群落结构的稳定性[35]。另外,Peco等[36]研究表明,草地禁牧后,由于物种间竞争平衡受家畜采食行为干扰程度降低,造成草地群落中少量竞争力较强的物种成为群落的优势种群,导致物种多样性下降。这与本研究在禁牧区内垂穗披碱草具有较高的重要值(表1),草地群落香农维纳指数、多样性指数、丰富度指数较低的结果一致。因此,合理的放牧制度能促进牧草生长,提高草地群落物种多样性。前人关于不同放牧制度对草地群落结构和物种多样性的研究中,禁牧、季节放牧及自由放牧区的差异均达到显著水平,但本研究中除季节放牧样区第二年香农维纳指数显著高于自由放牧区和禁牧区外,其余多样性指数差异均不显著,这可能是由于高原鼢鼠对草地的扰动,鼢鼠活动可能对草地群落多样性指数存在影响,其结果还需要进一步验证。
高原鼢鼠造丘及挖掘活动能影响草地群落结构和物种多样性,而高原鼢鼠对草地群落结构和物种多样性的影响与其食性有关。高原鼢鼠是广食性动物,影响其食性的因素主要与外界的因素有关,如栖息地气候因素、植物生长周期、食物的喜好性及可食植物与其他植物的种间种间种内竞争等[37]。王权业等[38]通过对鼢鼠胃内容物的显微结构分析发现,高原鼢鼠食物组成在不同栖息地、性别差异无关,而与不同植物出现的频次有关;高原鼢鼠主要取食杂类草根茎,主要取食植物为摩苓草(Morinachinensis)、美丽风毛菊、直立梗唐松草、丽江风毛菊(Saussurealikiangensis)、雪白委陵菜(Potentillanivea)、鹅绒委陵菜、棘豆、西伯利亚蓼(Polygonumsibiricum)、异叶米口袋和细叶亚菊(Ajaniatenuifolia),这与本研究中鹅绒委陵菜、美丽风毛菊、多裂委陵菜等杂类草在自由放牧区和季节放牧区具有较高重要值的研究结果相一致(表1);可见杂草类的种类和数量能在一定程度上反映该生境内的高原鼢鼠种群密度。高原鼢鼠种群数量增减与草地群落物种组成存在一定的联系,主要表现为:随着鼢鼠种群密度的增加,多年生禾本科牧草盖度减少,一年生或多年生的杂类草的盖度增加[23]。陶燕铎等[39]对高寒草甸草地防除杂草后发现,高原鼢鼠种群数量快速下降,从侧面证实杂草的数量和分布决定高原鼢鼠的种群数量和分布。同时,魏万红等[40]研究也发现,高原鼢鼠的种群密度与杂草类生物量和土壤硬度呈显著正相关。另外,张堰铭[24]等研究表明,植物群落物种丰富度与高原鼢鼠种群密度呈显著负相关,高原鼢鼠种群密度与鹅绒委陵菜、萼果香薷、细叶亚菊、西伯利亚蓼等高原鼢鼠喜食植物的数量有关,高原鼢鼠捕杀后单子叶植物重要值增加,鹅绒委陵菜、二裂委陵菜、萼果香薷、细叶亚菊等双子叶植物的重要值降低,且重要值降低的大多数物种与高原鼢鼠种群密度呈正相关关系。同时,本研究发现季节性放牧区的高原鼢鼠相对种群密度最高,而禁牧区相对种群密度最低;这与“中度干扰理论”的在中度扰动强度下,物种多样性最高、生态系统最稳定的假说一致。另外,禁牧区高原鼢鼠相对鼠群密度保持最低,这与单子叶植物地下生物量增加,阔叶类植物地下生物量下降,造成高原鼢鼠的食物不足等因素有关。
因此,放牧制度能影响草地群落各物种的重要值,改变高原鼢鼠喜食植物的组成和数量,进而改变高原鼢鼠的种群密度。高原鼢鼠危害的发生与草地载畜量相关,草地载畜量过高和高原鼢鼠的挖掘活动共同作用能加剧草地退化草地进程,这从自由放牧区种群数量较高、草地生物量较低、植物多样性指数较低等方面得到证实;而季节性放牧区虽然高原鼢鼠种群数量较高,但由于控制草食家畜的采食频次,草地群落结构及物种多样性保持最高。因此,在鼠害严重的区域早期通过禁牧措施,短期内实现草地优势种群-单子叶植物快速恢复,改善草地群落结构,减少高原鼢鼠的食物来源,进而降低高原鼢鼠种群密度;后期通过季节放牧,实现草地生物多样性和生态系统稳定的目标。
综上所述,季节性放牧可使草地群落物种数、草地建群种的重要值以及草地群落多样性指数都高于禁牧区和自由放牧区。草地群落地上生物量表现为:禁牧区>季节性放牧区>自由放牧;而地下生物量表现为:自由放牧区>禁牧区>季节性放牧区,且高原鼢鼠相对鼠群密度与不同放牧制度样地植物的地上、地下生物量存在相关性。因此,建立合理的放牧制度可以改善高寒草甸草地群落结构,进而调控高原鼢鼠的种群密度。
猜你喜欢
Zoological Research(2022年3期)2022-06-07
南方农业(2021年30期)2021-12-25
西藏艺术研究(2021年3期)2021-06-02
兽医导刊(2020年3期)2020-12-30
农家科技中旬版(2020年5期)2020-06-15
家教世界·创新阅读(2020年4期)2020-06-03
当代工人·精品C(2020年1期)2020-05-20
家教世界·创新阅读(2020年1期)2020-05-11
家教世界·创新阅读(2020年2期)2020-04-07
西藏农业科技(2019年1期)2019-07-25
