
乌拉尔图小麦PBF基因克隆及其序列分析
2018-08-01 07:53胡喜贵吴晓军姜小苓王玉泉胡铁柱茹振钢
江苏农业科学 2018年13期
胡喜贵, 吴晓军, 姜小苓, 王玉泉, 李 淦, 胡铁柱, 茹振钢
(河南省现代生物育种协同创新中心/河南科技学院小麦研究中心,河南新乡 453003)
DOF(DNA-binding with one finger)蛋白是植物特有的一类转录因子,在植物生长发育过程中起着重要作用[1]。DOF蛋白一般由200~400个氨基酸组成,包括2个主要结构域:N-端DNA结合高度保守结合域(DOF Domain)和C-端转录调控域[2]。醇溶蛋白盒结合因子(prolamin-box binding factor,PBF)属于DOF蛋白家族之一,是胚乳组织特异的基因表达调控因子,主要调控作物籽粒中贮藏蛋白基因表达,最终影响贮藏蛋白的积累量[3]。目前已在玉米、水稻、大麦等禾本科作物中,广泛开展了PBF基因调控功能研究[4-6]。Mena等利用大麦PBF基因同源性,首次克隆出小麦PBF编码基因[5]。该基因被定位于普通小麦(TriticumaestivumL.,AABBDD,2n=6x=42)第5号同源群(5A、5B和5D)的着丝粒附近,并在小麦胚乳的整个发育过程中持续表达[7]。
普通小麦是异源多倍体物种的一个典型代表,其A、B和D染色体组分别由乌拉尔图小麦(TriticumurartuTum. AA,2n=2x=14)、拟斯卑尓脱山羊草(AegilopsspeltoidesTausch. SS,2n=2x=14)和节节麦(AegilopsTauschiiCoss. DD,2n=2x=14)提供[8-12]。由于异源多倍化以及长期驯化的过程,现代普通小麦的遗传多样性较其祖先种大大降低[13]。Ravel等对27个普通小麦研究证实,普通小麦的PBF等位基因(wPBF-A、wPBF-B和wPBF-D)多样性较低[14]。因此,发掘普通小麦祖先种PBF基因的种质资源,将对小麦品质改良具有重要意义。截至目前,节节麦PBF基因已有报道[15],但其他2个祖先种的PBF基因尚未见报道。本研究拟对2份乌拉尔图小麦材料进行PBF基因克隆并分析其序列结构特征,以期为进一步研究乌拉尔图小麦PBF基因调控功能、改良普通小麦品质等奠定基础。
1 材料与方法
1.1 试验材料
本试验2份乌拉尔图小麦材料(TriticumurartuTum.)PI428225(土耳其)和PI428269(黎巴嫩),均由美国种质资源库(National Genetic Resources Program,NPGS)馈赠,经河南科技学院小麦中心繁殖并保存。
1.2 引物设计
根据NCBI数据库中乌拉尔图小麦G1812的全基因组鸟枪法测序数据(登录号:KD091395),设计1对覆盖PBF基因完整编码区的特异性引物:PBF-F(5′-ATTCGATATGTGT-GTACACATGT-3′)和PBF-R(5′-TCTTGCACTTACATCA-GGGAG-3′),其引物由Thermo Fisher Scientific InvitrogenTM公司合成。
1.3 基因克隆与测序
2份乌拉尔图小麦材料进行室温发苗,培养7 d左右,取幼嫩叶片备用。参照Yan等的2×CTAB方法[16],提取基因组总DNA。采用高保真TransStart®Fast Pfu DNA聚合物(TransGen Biotech,中国北京)进行PBF基因扩增。其PCR扩增程序:95 ℃ 5 min;95 ℃ 40 s,58 ℃ 30 s,72 ℃ 1 min,30个循环;72 ℃ 5 min,4 ℃保存。扩增产物用1.0%琼脂糖进行电泳分离,切取目的条带,经PUEX Gel DNA回收试剂盒(Bioche,中国北京)纯化,与pEASY克隆载体(pEASY®-Blunt Zero Cloning Kit,TransGen Biotech,中国北京)连接。经转化、筛选、挑取3个阳性克隆,送华大基因公司(BGI,中国深圳)测序。
1.4 序列分析
经测序分析,获得乌拉尓图小麦PBF基因DNA序列。利用NCBI的Blastn工具进行在线序列比对,判断其属性。同时,搜索与其同源的核苷酸及氨基酸序列。利用DNAMAN(Ver.7.0)进行多序列比对处理,DNASP(Ver.5.10.01)分析序列的多态性位点;利用NLStradamus(http://www.moseslab. csb.utoronto.ca/NLStradamus/)和MyHits (http://myhits.Isb-sib.ch/)分别进行蛋白核定位信号和翻译后修饰位点在线预测。运用MEGA Ver 4.0中的邻接法(neighbor joining method,NJ)构建聚类树。
2 结果与分析
2.1 乌拉尔图小麦PBF基因克隆
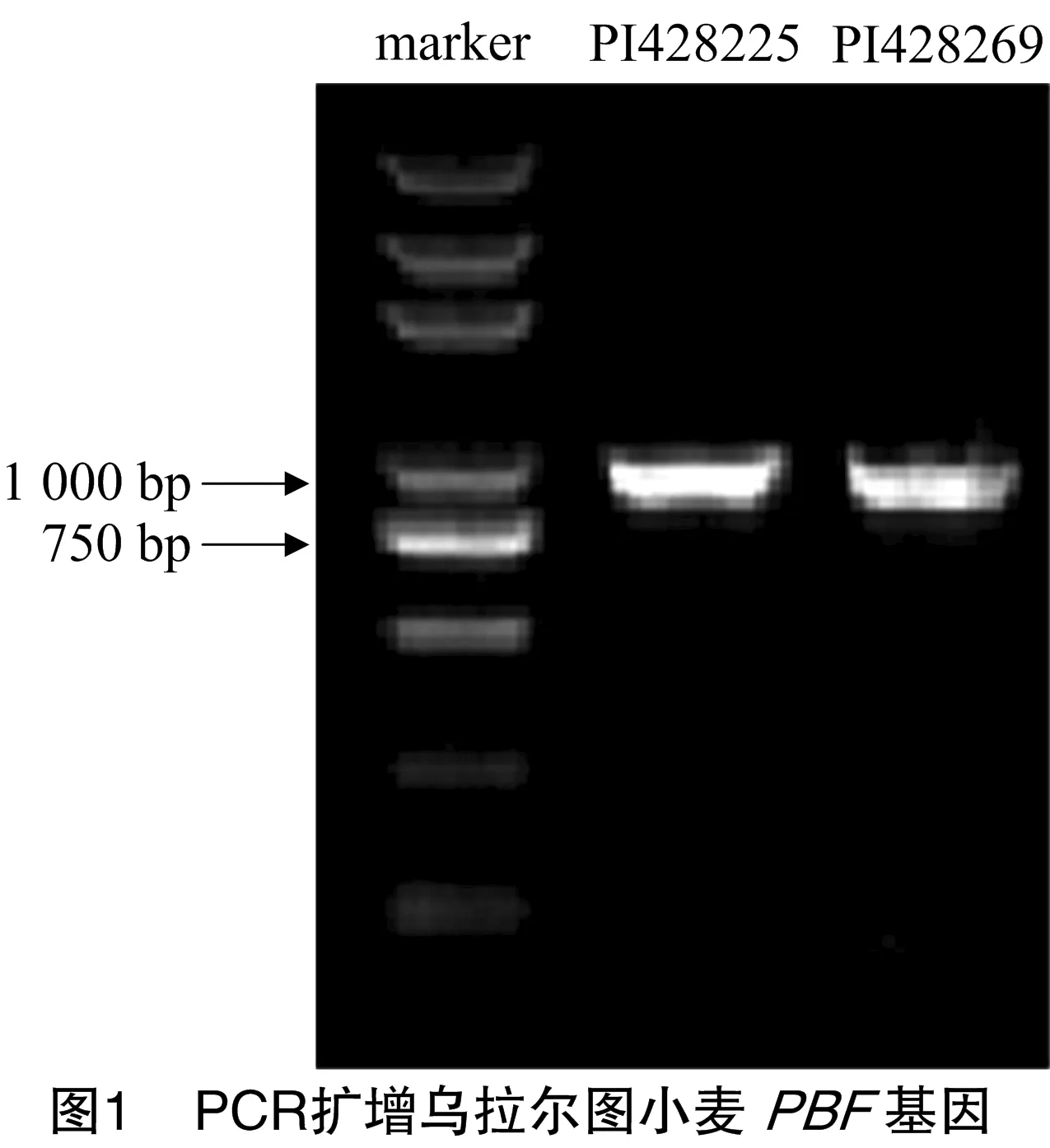
利用设计引物PBF-F和PBF-R对2份乌拉尔图小麦PI428225和PI428269的基因组DNA进行PCR扩增,其扩增产物经1.0%琼脂糖凝胶电泳,2份材料均扩增出长度为 1 000 bp 左右的单一条带(图1)。将目的片段回收、纯化,连接到pEASY克隆载体上,经测序分析后,2份材料的条带核苷酸序列长993 bp,对应GenBank数据库登录号为MF547409(PI428225)和MF547410(PI428269)。
2.2 乌拉尔图小麦PBF基因核酸序列比对分析
利用DNAMAN软件进行序列多重比对,表明,本研究克隆乌拉尔图小麦得到2个PBF基因的核苷酸序列(MF547409和MF547410),与已报道乌拉尔图小麦(EMS61761)、节节麦(KJ544771和KJ544772)和普通小麦中国春(KC849701、KC849702和KC849703)的PBF基因具有相同的长度(993 bp),一致性高达97.35%。但是,相比不同来源PBF基因序列分析发现,乌拉尔图小麦PBF存在丰富变异位点,高达45个单核苷酸多态性位点(SNPs)(图2)。在45个SNPs中,30个转换(transitions)位点包括18个G←→A(90、312、423、441、444、462、463、489、504、648、708、718、768、827、857、883、926、951)和12个C←→T(271、313、392、502、513、578、614、654、687、735、861、888),15个颠换(transversions)位点包括3个T←→A(14、573、590)、3个T←→G(64、635、800)和9个C←→G(68、285、336、366、713、794、891、911、980)。
对乌拉尔图小麦3个PBF基因的核苷酸序列比较分析发现,共存在6个突变位点。其中,已报道的EMS61761序列中有4个位点,分别是第59、第249位点的C→T、第393位点A→T、第514位点G→A;本研究的MF547410序列中仅存在2个位点,是第825、第938位点的C→T。可见,乌拉尔图小麦的PBF基因是相对保守的。
2.3 乌拉尔图小麦PBF基因氨基酸序列比对分析
对根据乌拉尔图小麦、节节麦和普通小麦中国春的PBF基因推导的氨基酸序列进行比较发现,它们具有相同长度330个氨基酸(aa),并具有相似的结构特征,均含有蛋白核定位信号保守序列KKPR(NLS core),N-端保守的DOF结构域(DOF domain),C-端可变的调控区域具有典型的富天冬酰胺(asparagine-rich)序列特征,以及连接N-端结构域和 C-端调控域的丝氨酸残基链(ser hinge)(图3)。但相比节节麦、普通小麦中国春的PBF氨基酸序列,乌拉尔图小麦PBF氨基酸序列发现22个氨基酸变异位点(图3),主要归因于核苷酸序列上碱基的替换(转换和颠换)(图2)。其中,7个变异位点(5、22、23、91、95、105、131)分布于N-端DOF结构域之外;15个变异(155、193、197、205、212、238、240、265、267、276、286、295、304、309、327)分布于C-端调控区域。
MyHits分析PBF氨基酸序列,共存在3种类型的修饰位点,其中,N-端糖基化位点(ASN-glycosylation site)均有4个,1个位于N-端保守的DOF结构域,其余3个集中分布于C-端调控区域末端;酪氨酸激酶Ⅱ磷酸化位点(CK2-phospho-site)均有5~6个,分布于整条PBF氨基酸序列上;蛋白激酶C磷酸化位点(PKC-phospho-site)均有2个,分别位于N-端保守的DOF结构域和C-端富天冬酰胺区域。进一步分析发现,乌拉尔图小麦PBF氨基酸序列存在特有1个酪氨酸激酶Ⅱ磷酸化位点(CK2-phospho-site),该位点因第22、第23位点2个氨基酸变异(A→S和G→A)形成,这一特征显著区别其他来源序列(图3)。
2.4 聚类分析
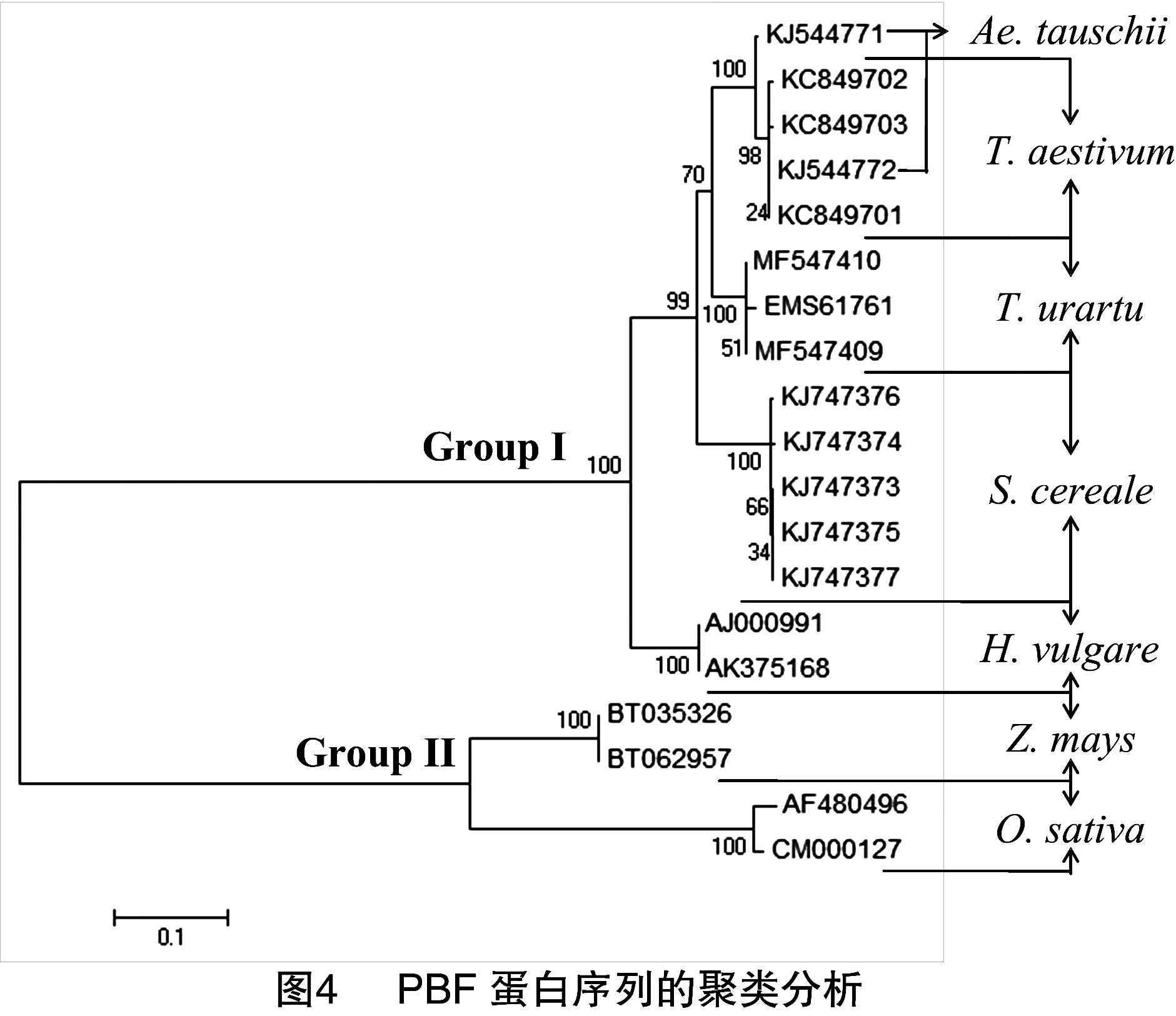
NCBI数据库中有17个PBF序列:普通小麦中国春,KC849701、KC849702和KC849703;节节麦,KJ544771和KJ544772;乌拉尔图小麦,EMS61761;黑麦(Secalecereale),KJ747373~KJ747377;大麦(Hordeumvulgare),AJ000991和AK375168;玉米(Zeamays),BT035326和BT062957;水稻(Oryzasativa),AF480496和CM000127和本研究的2个乌拉尔图小麦PBF序列构建的聚类树(图4),19个PBF序列被清晰地分成2组(GroupⅠ和Group Ⅱ)。在第1组(Group Ⅰ)中,15个PBF序列被进一步分为3个亚组,即来源于大麦PBF亚组、来源于黑麦PBF亚组、来源于普通小麦及供体祖先种(节节麦和乌拉尔图小麦)PBF亚组。
3 讨论与结论
Ravelc等对27个普通小麦PBF基因研究发现,wPBF-A、wPBF-B、wPBF-D等位基因分别存在1、5、1个单核苷酸多态性位点(SNPs),其结果表明普通小麦中PBF基因遗传多样性较低[14]。本研究从2份乌拉尔图小麦材料克隆出PBF基因(MF547409和MF547410)核苷酸序列与NCBI数据库中乌拉尔图小麦PBF基因序列(EMS61761)具有较高一致性,仅存在6个突变位点。然而,相比节节麦和普通小麦中国春PBF基因, 乌拉尔图小麦PBF存在丰富变异位点,高达45个单核苷酸多态性位点(SNPs)(图2),表明乌拉尔图小麦PBF基因具有较高遗传多样性。聚类分析显示,来源于乌拉尔图小麦与普通小麦的PBF聚在1个亚组(图4),证实了二者亲缘关系较近[10]。同时,又进一步印证了PBF蛋白在物种间存差异且亲缘关系越远,其差异越大,即PBF蛋白表现为种属特异性[15]。

DOF是植物特有的转录因子家族,典型的DOF蛋白由200~400个氨基酸组成,其N-端有高度保守的DOF结构域和C-端转录调控域[1-2]。DOF蛋白能够特异地识别并结合真核生物基因启动子区域的顺式作用元件,如CCN4-like(GLM)、醇溶蛋白盒(prolamin box,PB)、5′-AACA/TA-3′基序等,从而激活或抑制基因转录[17]。DOF结构域外的氨基酸对识别能力没有影响,但核心序列之外的侧翼序列对DOF蛋白与DNA结合特性有一定的影响[18]。C-端转录调控域可接受不同信号途径的调控,与不同类型调控蛋白相互作用激活或抑制基因的转录[1]。可见,DOF蛋白中氨基酸序列变异可导致其功能多样性。已有研究表明,普通小麦wPBF与小麦醇溶蛋白、麦谷蛋白中的顺式作用元件PB特异结合,参与这2类贮藏蛋白基因的表达调控[19-20]。本研究推导的乌拉尔图小麦PBF蛋白与来源节节麦、普通小麦中国春PBF蛋白序列对比发现,所有PBF蛋白均具有DOF典型结构特征。然而,乌拉尔图小麦PBF蛋白序列存在22个氨基酸变异位点:7个位于N-端DOF结构域之外和15个位于C-端调控区域,其中2个氨基酸变异(第22位点,A→S;第23位点,G→A)形成了乌拉尔图小麦特有酪氨酸激酶Ⅱ磷酸化位点。这些氨基酸变异位点可能对乌拉尔图小麦PBF蛋白的调控功能产生影响,仍须进一步功能研究证实。综上所述,本研究为进一步研究乌拉尔图小麦PBF基因的相关特性、后续特异标记开发、功能分析等奠定基础。

猜你喜欢
华北农学报(2021年1期)2021-03-26
广州大学学报(自然科学版)(2019年1期)2019-05-07
西南农业学报(2018年9期)2018-10-19
新疆农垦科技(2016年2期)2016-08-21
现代兵器(2016年4期)2016-04-27
天津科技大学学报(2016年1期)2016-02-28
山东农业科学(2015年11期)2016-01-11
湖北师范大学学报(自然科学版)(2015年2期)2016-01-10
现代检验医学杂志(2015年2期)2015-02-06
植物营养与肥料学报(2011年6期)2011-10-24
