
内分泌干扰物对胰腺发育的影响及其机制研究进展
2018-08-01 01:18王洪芳赵飞王蔚汝少国
生态毒理学报 2018年3期
王洪芳,赵飞,王蔚,汝少国
中国海洋大学海洋生命学院,青岛 266003
1 胰腺发育的调控机制(Regulating mechanisms of pancreas development)
胰腺是由内分泌腺和外分泌腺组成的双功能腺体,其中外分泌腺由腺泡细胞和导管细胞组成,主要功能是分泌消化酶进入肠道对食物进行消化;而内分泌腺由胰岛组成,主要功能是分泌激素维持血糖相对恒定。胰岛包含α、β、δ、ε和PP细胞5种内分泌细胞,各种细胞通过分泌不同的激素维持血糖平衡。其中β细胞占胰岛的绝大部分,分泌胰岛素,降低血糖;α细胞分泌胰高血糖素,升高血糖。因此,内分泌胰腺功能紊乱会导致葡萄糖稳态受损,或引发糖尿病[8-11]。
脊椎动物的胰腺由前肠内胚层上皮的背侧和腹侧外翻部分发育而来[10-11]。在小鼠中,胚胎发育9.5 d(9.5 days of gestation, e9.5),胰腺以2个独立胰芽(背芽和腹芽)的形式出现。早期胰芽(e9.5~12.5)主要由多能胰腺祖细胞组成,该时期为胰腺祖细胞活跃增殖期。e11.5~12.5,2个胰芽融合在一起。e12.5,胰腺上皮外翻侵入邻近的间质,随后进行扩张和分支。e13.5,位于分支尖端的多能胰腺祖细胞分化为腺泡祖细胞,而分支主干上的多能胰腺祖细胞成为双潜能祖细胞,并分化出内分泌祖细胞和导管细胞。e14.5,腺泡祖细胞分化成腺泡细胞,内分泌祖细胞移离主干上皮形成内分泌细胞簇。在出生后的发育过程中,成熟的内分泌细胞聚集形成胰岛,产生了具有腺泡、导管和胰岛的成熟胰腺[8,10-11]。
胰腺的发育由转录因子和外周信号分子相互作用形成的复杂的基因调控网络协调调控。谱系追踪研究表明,转录因子是调控胰腺器官发生、分化和成熟的关键因素,在胰腺发育的每个阶段都有许多转录因子表达[8](见图1)。按照胰腺发育顺序,可将转录因子分为以下几种类型:Pdx1、Ptf1a、Mnx1、Sox9、Nkx6.1、HNF1β、HNF6、Foxa2和GATA4/6参与多能胰腺祖细胞的形成和增殖;Pdx1、Ptf1a、Sox9、Nkx6.1、HNF1β和GATA4决定分支尖端多能胰腺祖细胞的形成;Pdx1、Sox9、Nkx6.1、HNF1β、HNF6、Foxa2和GATA6参与分支主干上双潜能祖细胞的形成;Ptf1a和GATA4使分支尖端的多能胰腺祖细胞定向分化为腺泡细胞;HNF1β、HNF6、Sox9和Foxa2等调控分支主干的双潜能祖细胞分化为导管细胞;Ngn3及其直接靶基因Islet1和NeuroD1决定双潜能祖细胞定向分化为内分泌祖细胞;最后,转录因子Mnx1、Islet1、NeuroD1、Pdx1、Pax4/6、Nkx2.2/6.1和MafA等决定内分泌祖细胞向何种内分泌细胞分化[8,10-11]。除了胰腺转录因子的级联调节,非胰腺组织的外部信号对胰腺发育也至关重要。其中Notch信号抑制多能胰腺祖细胞分化直至时机成熟,调控内分泌祖细胞分化为β细胞。FGF信号参与胰芽形成以及促进多能胰腺祖细胞增殖。Wnt信号参与腹芽形成,促进多能胰腺祖细胞增殖,并调控胰腺内外分泌细胞发育和β细胞功能。RA信号在内胚层形成、背芽发育和多能胰腺祖细胞分化中起着重要作用。在胰腺发育的早期阶段,Hedgehog信号必须被抑制才能保证胰芽正常发育。TGFβ信号在胰芽形成中必不可少。VEGF信号参与背芽发育;BMP信号参与腹芽的形成[8,10,12-13]。此外表观遗传机制也可能在胰腺发育中发挥重要作用:研究证实前肠内胚层中的组蛋白甲基转移酶缺失会导致腹芽形成受阻,转而发育为肝芽,表明组蛋白修饰决定了肝脏和腹芽的命运[10]。
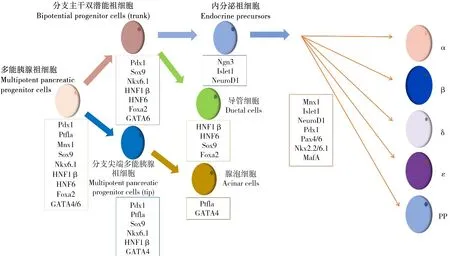
图1 胰腺发育及相关转录因子Fig. 1 Pancreas development and the related transcription factors
2 EDCs对脊椎动物胰腺发育的影响(Effects of endocrine disrupting chemicals on pancreas development of vertebrates)
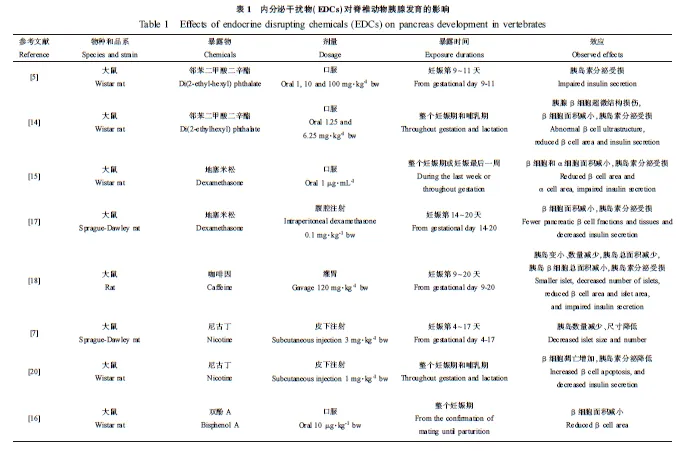
根据物种、EDCs的种类、剂量、暴露时间和效应,将EDCs对胰腺发育的影响的研究结果进行整理,见表1。
2.1 哺乳动物
动物模型实验研究表明,EDCs主要对大鼠的内分泌胰腺发育具有干扰和破坏作用,表现为以下3个方面。
胰岛β细胞结构损伤。研究发现Wistar大鼠在妊娠期和哺乳期暴露于邻苯二甲酸二辛酯,导致子代断乳后胰岛β细胞超微结构损伤[14]。
胰岛尺寸降低。邻苯二甲酸二辛酯能够降低发育期Wistar大鼠的胰岛β细胞面积[14]。孕期Wistar大鼠暴露于地塞米松,结果胚胎的β细胞面积和α细胞面积都减小[15]。Wistar大鼠暴露于双酚A,仔鼠出生时的β细胞面积降低[16]。研究发现大鼠产前暴露于地塞米松,也导致子代β细胞面积减小[17]。孕期咖啡因暴露导致胚胎胰岛变小、数量减少,胰岛总面积减少,胰岛β细胞总面积减小[18]。Sprague-Dawley大鼠孕期暴露于尼古丁,结果子代胰岛数量减少、尺寸降低,进而导致内分泌胰腺在整个胰腺中的比例显著降低[11]。Wistar大鼠妊娠期暴露于盐酸舍曲林,导致子代胰岛β细胞面积减小[19]。Wistar大鼠在妊娠期和哺乳期暴露于尼古丁,结果仔鼠出生时β细胞凋亡增加,β细胞面积减小[20]。
胰岛β细胞功能障碍(胰岛素分泌受损)。大鼠妊娠期暴露于邻苯二甲酸二辛酯会损伤发育期子代胰岛素的分泌,降低胰腺中的胰岛素含量[9,14]。孕鼠在妊娠最后一周和整个妊娠期暴露于地塞米松,造成子代胰岛素分泌缺陷[15]。大鼠产前暴露于地塞米松导致仔鼠胰岛素水平降低[17]。孕鼠孕期暴露于咖啡因导致子代β细胞内胰岛素分泌颗粒数目减少,囊泡内颗粒的膜融合和脱落现象明显[18]。尼古丁染毒雌性Wistar大鼠导致子代胰岛素分泌降低[20]。
宝格丽Octo Finissimo陀飞轮自动上链腕表厚度仅为3.95毫米,第四次刷新世界纪录。Octo Finissimo陀飞轮自动上链腕表作为2018年又一具有里程碑意义的新作,充分诠释了宝格丽在腕间时计上所赋予的非凡艺术造诣。Octo Finissimo镂空陀飞轮腕表与Octo Finissimo自动上链腕表均在去年的日内瓦高级钟表大赏斩获大奖,为品牌赢得业内一致认可。
2.2 其他动物
胰腺的发生过程以及调控胰腺发育的转录因子和信号通路在脊椎动物中高度保守,因此除哺乳动物外,斑马鱼等其他脊椎动物也被广泛用于胰腺发育及功能的相关研究。斑马鱼具有胚体透明、体外发育迅速、养殖成本低等优点,针对胰腺特异性基因开发的转基因技术使斑马鱼胚胎成为研究外源污染物对胰腺发育影响的有力体内实验模型。例如,Tg(ins:GFP)转基因株系可以在生成胰岛素的β细胞中表达绿色荧光,使胰岛可见;Tg(ptf1a:GFP)株系可以在外分泌胰腺组织中表达绿色荧光,使外分泌胰腺可见[21]。利用转基因斑马鱼胚胎,目前的研究发现EDCs对内分泌胰腺和外分泌胰腺发育的影响主要包括以下4个方面:
胰岛面积减小。将受精后3 h(3 hours postfertilization, 3 hpf)的Tg(ins:GFP)和Tg(gcga:GFP)转基因斑马鱼胚胎暴露于200 μg·L-1邻苯二甲酸异辛酯,结果斑马鱼胚胎和仔鱼的β细胞和α细胞的面积显著减小[22]。全氟辛基磺酸染毒Tg(ins:GFP)和Tg(ptf1a:GFP)转基因斑马鱼胚胎,导致斑马鱼胚胎的初级胰岛面积减小[23]。
胰岛畸形。邻苯二甲酸异辛酯暴露增加了斑马鱼胚胎和仔鱼胰岛形态变异的频率[22]。全氟辛基磺酸暴露导致斑马鱼胚胎和仔鱼胰岛形态发生变异的频率增加[23]。将Tg(ins:GFP)和Tg(ins:荧光探针)转基因斑马鱼胚胎暴露于3,3’,4,4’,5-五氯联苯,结果发现3,3’,4,4’,5-五氯联苯导致胰岛形态变异的频率增加,初级胰岛过早地迁移,胰岛破碎以及形成异位β细胞[24]。研究发现,曲古抑菌素A和丙戊酸可以通过破坏斑马鱼胰岛内分泌细胞的聚集扰乱斑马鱼内分泌胰腺的发育[25]。次级胰岛形成延迟。全氟辛基磺酸能够延迟Tg(ins:GFP)和Tg(ptf1a:GFP)转基因斑马鱼次级胰岛的形成[23]。
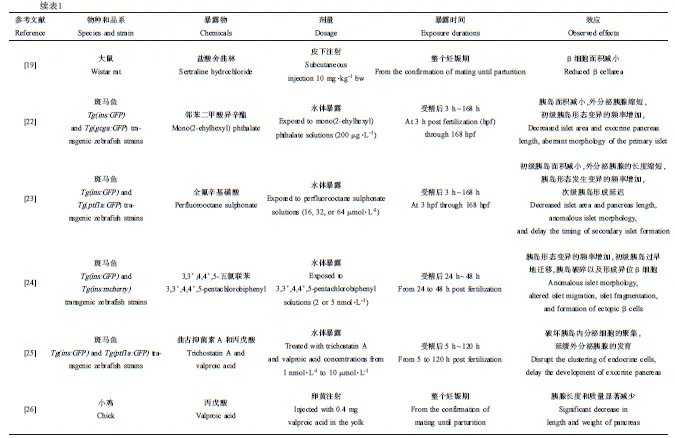
外分泌胰腺长度缩短。将Tg(ptf1a:GFP)转基因斑马鱼胚胎暴露于邻苯二甲酸异辛酯,导致外分泌胰腺占鱼身体总长的比例显著降低,外分泌胰腺缩短[22]。全氟辛基磺酸和邻苯二甲酸异辛酯类似,也缩短了外分泌胰腺的长度[23]。曲古抑菌素A和丙戊酸可以延缓斑马鱼外分泌胰腺的发育[25]。另外,Akhtar等[26]研究发现,丙戊酸暴露导致小鸡胚胎的外分泌胰腺长度显著减少。
3 机制研究(Study on related mechanisms)
3.1 影响转录因子的表达
各种胰腺转录因子参与了胰腺发育和β细胞分化。在这些转录因子中Pdx1主要负责调控胰芽形成、β细胞分化以及维持成熟β细胞的功能;Pdx1可以通过结合胰岛素基因转录调控区A3激活其转录过程,上调胰岛素表达水平,是胰腺发育的主要调控因子[27-29]。Ngn3是胚胎时期决定胰腺内分泌细胞分化的重要因子,谱系追踪试验表明所有的内分泌细胞都来源于表达Ngn3的祖细胞[11]。胰腺形成过程中的另一个关键转录因子Ptf1a主要负责调控外分泌细胞的分化以及维持外分泌细胞的功能[11,30]。Nkx2.2和Nkx6.1属同源域蛋白,Nkx2.2对维持内分泌细胞的数量起重要作用,敲除小鼠的Nkx2.2导致β细胞完全缺失,α和γ细胞的数量急剧减少,ε细胞的数量增加;Nkx6.1仅在β细胞中表达,对β细胞的生成起重要作用,Nkx6.1基因缺失的小鼠无法产生β细胞。Pax4和Pax6也属同源域转录因子,Pax4功能受损或缺失,小鼠的β细胞和δ细胞缺失,α细胞增多;Pax6基因突变会导致α细胞缺失,β细胞、δ细胞和γ细胞数量减少。NeuroD1在胰岛内分泌细胞的分化过程中起重要作用,NeuroD1基因失活的小鼠,在e14.5~17.5胰岛发育停滞,小鼠出生后出现严重的糖尿病[11,13]。MafA是β细胞特异性转录因子,是胰岛素基因转录的有效激活剂[27,31]。HNF4α是β细胞转录的全局性调控因子[32]。
研究发现,环境中很多EDCs可以通过影响胰腺发育中关键转录因子的表达,对胰腺器官的形成造成干扰或损伤。在邻苯二甲酸二辛酯对发育期大鼠胰腺发育的影响研究中发现,大鼠妊娠期暴露于邻苯二甲酸二辛酯可降低子代内分泌胰腺发育关键转录因子Pdx1、Pax4、Pax6、MafA和HNF4α的表达,造成胰岛β细胞损伤,包括超微结构损伤、面积减小、胰岛素分泌受损,胰腺中的胰岛素含量降低[5,14]。胚胎期暴露于双酚A可降低出生时仔鼠胰腺发育关键转录因子Pdx1的表达,从而影响β细胞的正常发育,降低β细胞的质量[16]。地塞米松可通过降低子代大鼠Pdx1、Ngn3、Pax6、NeuroD1和MafA的表达导致β细胞和α细胞面积减小,胰岛素分泌受损,胰岛素含量降低[14,17]。咖啡因暴露后内分泌发育相关转录因子Pdx1、Ngn3、NeuroD1、Nkx6.1和Pax6的表达均显著降低,导致内分泌细胞发育受损,胰岛尺寸降低、数量减少,β细胞面积减小,胰岛素分泌被破坏[18]。尼古丁可下调发育期大鼠Pdx1、Pax6和Nkx6.1的表达,导致胰岛尺寸降低,胰岛数量减少,使内分泌胰腺在整个胰腺中的比例显著降低[7]。盐酸曲舍林暴露会降低子代大鼠Pdx1、Ngn3和NeuroD1的表达,导致β细胞面积减小[19]。斑马鱼胚胎暴露于邻苯二甲酸异辛酯显著下调内分泌胰腺发育关键转录因子Pdx1的表达,导致β细胞和α细胞的面积显著减小,即胰岛面积减小,胰岛形态变异的频率增加;外分泌胰腺发育关键转录因子ptf1a的表达显著降低,导致外分泌胰腺长度缩短[22]。斑马鱼胚胎暴露于全氟辛基磺酸显著降低了Pdx1的表达,导致初级胰岛面积减小,胰岛形态发生变异的频率增加,次级胰岛形成延迟[23]。小鸡胚胎暴露于丙戊酸会干扰内分泌胰腺的生长发育,可能是由于抑制了内分泌胰腺发育关键转录因子Pax6的表达使胰岛的正常发育受损[26]。
此外,研究发现性激素与其受体结合后,能够调控胰腺发育关键转录因子的表达,进而影响胰腺的发育过程。Rae等[33]研究报道,孕期雄激素(丙酸睾酮)能通过结合雄激素受体上调转录因子Pdx1的表达,增加β细胞数量;Yuchi等[34]研究报道,17β-雌二醇能够通过结合雌激素受体α(estrogen receptor α, ERα)调控Ngn3的表达,进而促进胰腺发育过程中β细胞的增殖和分化。这些研究表明性激素对胰腺发育也发挥调控作用,而环境中很多EDCs能够竞争性结合雌/雄激素受体而发挥抗雌/雄激素效应,如雌马酚、染料木黄酮、三苯氧胺等能竞争结合雌激素受体、而甲氧滴滴涕、氟他胺、甲基睾酮、乙烯菌核利代谢物等能竞争结合雄激素受体[35];因此,这类EDCs可能会通过竞争结合性激素受体的途径最终改变胰腺发育相关转录因子的表达水平,但相关研究尚未见报道,还需进一步探讨。
3.2 诱导氧化应激
研究表明,许多内分泌干扰物可以通过诱导氧化应激造成胰腺β细胞损伤。氧化应激是由于活性氧自由基(reactive oxygen species, ROS)产生过量造成的,由于胰腺β细胞中抗氧化酶的表达水平较低,因此很容易受到氧化应激的影响。ROS可以通过多种机制影响β细胞的功能和存活,包括改变酶活性、离子通道运输和受体信号转导,诱发基因表达的异常调控以及细胞凋亡。
研究发现,地塞米松暴露妊娠期Sprague-Dawley大鼠增加了胎鼠胰腺中氧化应激标志物8-羟基-2-脱氧鸟嘌呤核苷的含量,表明地塞米松诱发了氧化应激,并最终破坏胰腺发育,使β细胞面积减小,胰岛素分泌受损[17]。Jacobs等[22]研究证实了氧化应激参与了邻苯二甲酸异辛酯对Tg(ins:GFP)和Tg(gcga:GFP)转基因斑马鱼胰腺器官发生的破坏:邻苯二甲酸异辛酯暴露后胰腺细胞内抗氧化物质谷胱甘肽(GSH)被氧化,该结果表明邻苯二甲酸异辛酯破坏了细胞的氧化还原环境,进而导致胰腺β细胞面积减小。此外,研究发现胰腺β细胞暴露于邻苯二甲酸二辛酯可以诱导ROS产生,导致GSH耗竭,破坏抗氧化酶超氧化物歧化酶(SOD)的活性,并导致丙二醛(MDA)积累,表明邻苯二甲酸二辛酯可能造成β细胞氧化损伤[36]。Bruin等[37]研究发现,Wistar孕鼠暴露于尼古丁能通过尼古丁乙酰胆碱受体在胚胎和新生大鼠发育过程中诱导胰腺产生氧化应激:显著增加了胰腺中谷胱甘肽过氧化物酶(GPx)和锰超氧化物歧化酶(SOD2)的蛋白表达,以及ROS的生成,进而介导β细胞凋亡。
研究发现转录因子Pdx1是氧化应激损伤胰腺发育的重要靶点:氧化应激发生时,ROS激活β细胞中的应激激活蛋白激酶JNK,促使转录因子Pdx1从细胞核转移到细胞质中,导致Pdx1的核积累减少,DNA结合活性降低,从而降低胰岛素基因的表达,最终损伤胰岛素的合成和分泌[38-40]。De Long等[19]报道Wistar大鼠妊娠期暴露于盐酸曲舍林后,子代胰腺发生氧化应激,损伤了胰腺关键转录因子Pdx1的表达,导致β细胞的面积减少;然而,EDCs诱发β细胞氧化应激后,是否通过激活JNK信号通路损伤Pdx1的表达,尚不清楚。
3.3 改变表观遗传修饰
表观遗传是指基因的序列不发生变化,但基因的表达发生了可遗传和可恢复的改变[41-42]。研究发现,在发育的关键时期(产前和产后)暴露于EDCs可以导致胰腺发育关键基因的表观遗传修饰异常,进而影响其正常表达、破坏胰腺发育。
3.3.1 影响DNA甲基化
DNA甲基化主要指在DNA甲基转移酶(DNA methyltransferase, DNMT)的催化下,胞嘧啶嘧啶环5’位置上加入一个甲基,主要发生在CpG双核苷酸的胞嘧啶碱基上;CpG双核苷酸在基因启动子区常聚集形成CpG岛,启动子区CpG岛的高甲基化和转录沉默相关[43-44]。研究发现,妊娠期Wistar大鼠暴露于邻苯二甲酸二辛酯后,子代胰岛中DNA甲基转移酶(Dnmt1, Dnmt3a和Dnmt3b)的表达水平升高,整体DNA甲基化水平升高,这表明邻苯二甲酸二辛酯暴露可能引发胰岛发育相关基因启动子区的高甲基化;甲基化DNA结合蛋白MeCP2和MBD2的基因表达也升高,推测甲基化的启动子区可能和MeCP2、MBD2共同作用,通过招募组蛋白乙酰化酶和其他因子,阻碍转录因子与DNA序列的结合,从而抑制β细胞发育和功能相关基因的表达,并最终导致β细胞功能障碍[5,45]。
环境雌激素如双酚A、4-壬基酚、乙烯雌酚等均能与雌激素受体(estrogen receptors, ERs)结合[35],而最近的研究发现ERs可能通过作用于DNMT等改变基因DNA甲基化水平。Marques等[46]报道乳腺癌中ERα可以通过招募Dnmt3B至基因启动子区,增加启动子的DNA甲基化水平,从而抑制基因表达;而Rüegg等[47]研究发现ERβ可能通过招募胸腺嘧啶糖苷酶、或阻止DNMT与启动子结合从而降低基因启动子区甲基化水平。这些研究提示EDCs特别是环境雌激素可能会通过结合ERs、调控DNA甲基转移酶等的功能改变基因启动子区甲基化水平,从而调控基因表达。然而,尽管研究发现胰腺细胞中也大量表达雌激素受体[48],且Pdx1[49]和胰岛素[50]等很多胰腺发育和功能相关基因的表达也受DNA甲基化调控,但EDCs对胰腺发育和功能相关基因表达的影响是否与其激活雌激素受体—DNA甲基转移酶途径相关,目前尚不清楚,还需进一步研究。
3.3.2 影响组蛋白的修饰
组蛋白修饰包括乙酰化、甲基化、磷酸化、泛素化、类泛素化和ADP-核糖基化修饰,这些修饰会影响核小体中的DNA与转录复合物的结合[51],继而影响基因的表达。组蛋白修饰也参与调控胰岛素基因的表达和胰腺的发育:研究发现与其他细胞相比,胰岛β细胞中胰岛素基因启动子邻近区域的组蛋白3(H3)乙酰化水平以及组蛋白3赖氨酸4(H3K4)甲基化水平均明显升高[52];Haumaitre等[53]使用组蛋白去乙酰化酶抑制剂处理大鼠胰腺细胞,发现组蛋白乙酰化能够影响外分泌胰腺中腺泡和导管的分化、促进内分泌胰腺的分化。研究表明,妊娠期Wistar大鼠暴露于双酚A可以改变子代胰腺发育关键调控基因Pdxl启动子区域的组蛋白修饰:子代胰腺Pdxl启动子区的组蛋白H3和H4乙酰化降低,组蛋白H3第4位赖氨酸三甲基化降低、第9位赖氨酸双甲基化升高,这些组蛋白修饰的变化可能抑制了Pdxl表达,进而损伤胰腺β细胞的发育,降低仔鼠出生时的β细胞面积[16]。
4 结语(Conclusions)
胰腺发育异常会导致糖尿病、肥胖、胰腺癌等多种疾病。环境中存在的EDCs可以通过不同的作用机制干扰脊椎动物生命早期的正常胰腺发育,尤其是内分泌胰腺发育,增加成年期患糖尿病、肥胖等代谢疾病的风险。本研究揭示了环境中常见的EDCs对胰腺发育的影响及其作用机制,为人体健康风险评估提供参考。
猜你喜欢
医学信息(2022年9期)2022-11-27
实用器官移植电子杂志(2022年5期)2022-03-24
小天使·二年级语数英综合(2021年8期)2021-08-16
中成药(2017年6期)2017-06-13
中国组织化学与细胞化学杂志(2016年4期)2016-02-27
安徽医科大学学报(2015年9期)2015-12-16
医学研究杂志(2015年11期)2015-06-10
肝胆胰外科杂志(2015年1期)2015-02-27
中国医学科学院学报(2014年6期)2014-03-11
中国海洋大学学报(自然科学版)(2014年6期)2014-02-28
