
根区温度对黄瓜嫁接苗碳氮代谢及相关酶活性的影响
2018-08-10 07:15孙世君付崇毅崔世茂
西北农林科技大学学报(自然科学版) 2018年8期
孙世君,付崇毅,崔世茂,宋 阳,高 宇,刘 琦
(1 内蒙古农业大学 农学院,内蒙古 呼和浩特 010019;2 内蒙古农牧业科学研究院,内蒙古 呼和浩特 010031)
利用日光温室和塑料大棚进行黄瓜冬季栽培和早春育苗的过程中,经常会受到低温胁迫的危害,使得植株丧失正常的生理功能[1]。针对生产中遇到的低温胁迫问题,前人研究主要集中在低气温胁迫方面。低气温会破坏植物体内的各种生理机制,抑制叶绿素的形成,降低植物光合作用,使植物的生长发育受到影响[2-3]。然而,事实上低地温对植物生长发育的影响也很大。低地温不仅影响根系对水分和矿质营养的吸收以及根系向地上部输送激素的含量[4],而且通过改变叶片中叶绿素含量、气孔阻力、内部酶活性等导致了光合产物和毒物的积累,进而影响了光合作用[5-6]。由此可见,根区温度对植物的生长起到重要的作用。
黄瓜在我国北方设施温室中的栽培比较广泛,然而由于黄瓜具有喜温不耐冷的特性,因此也常常因为受到不适宜根区温度的影响而不能正常生长发育。近几年来,有关根区温度对黄瓜生长影响方面的研究逐渐增多,但多集中在根区温度对黄瓜形态指标[7-8]、抗逆性指标[9-10]、矿质元素吸收[11-12]的影响等方面,而对其生命活动中最重要的碳氮代谢及相关酶活性方面的研究则相对较少,因此本试验以此为突破点,分析根区温度对黄瓜生长发育的影响,以期为冬季黄瓜生产提供理论依据。
1 材料与方法
1.1 试验材料与处理
1.1.1 试验材料 试验采用的黄瓜品种为津优35号,以黄瓜自根苗和嫁接苗为处理材料,嫁接苗的砧木包括黑籽南瓜和白籽南瓜,本研究以黑籽南瓜为砧木嫁接的黄瓜苗标记为“黑籽”,以白籽南瓜为砧木嫁接的黄瓜苗标记为“白籽”,自根黄瓜苗标记为“自根”。
1.1.2 试验处理 试验于2016年4-6月在内蒙古农业大学教学基地的日光温室内进行。4月3日开始播种砧木白籽南瓜,4月6日播种砧木黑籽南瓜,4月10日播种接穗,4月18日进行嫁接,5月6日当嫁接苗和自根苗均长至2叶1心时,挑选生长整齐一致的嫁接苗和自根苗开始进行温度处理。试验将根区温度设适温(18~20 ℃,CK)、亚适温(13~15 ℃)、低温(8~10 ℃)3个处理,每天处理8 h(22:00-06:00)。处理20 d后,取植株鲜样用液氮固定后超低温保存,用于各项生理指标的测定。
1.1.3 根区温度处理装置 该装置整体呈扁箱体,由高密度聚笨板包裹而成,扁箱体上方按照所需株行距打孔定植黄瓜苗。扁箱体内盛放土壤,在土壤层下方设置盘制循环冷水管,所述盘制循环冷水管一端与水泵连接后插入到冰柜底部作为出水管,另一端同样插入冰柜作为回水管,冰柜的上盖用同样大小的高密度聚苯板代替。在冷水管间隙设置盘制地热线,水泵和地热线各自连接一个温控器,并以并联的方式连接到时控器上,温控器的探头均放在扁箱体内底部中心位置。时控器通过控制2个温控器,从而自动控制某段时间内的土壤温度。具体结构如图1所示。

1.扁箱体;2.定植孔;3.地热线;4.冷凝管;5.温控器1探头;6.温控器1;7.水泵;8.冰柜;9.温控器2探头;10.温控器2;11.时控器;12.总电源1.Flat box;2.Planting hole;3.Ground heat pipe;4.Condenser tube;5.Temperature controller 1 probe;6.Temperature controller 1;7.Water pump;8.Freezer;9.Temperature controller 2 probe;10.Temperature controller 2;11.Timer;12.Total power supply图1 根区温度控制装置立体示意图Fig.1 Root-zone temperature control device
1.2 测定项目与方法
1.2.1 根区温度和气温 5月6日至5月26日,每隔5 d测定1次夜间(22:00-06:00)根区温度和气温,其中气温测定方法为:将温度记录仪(精创RC-4)探头置于各处理上方10 cm处,每30 min自动记录1次;根区温度用温度记录仪测定,土层深度为0~5 cm,每30 min自动记录1次。
1.2.2 黄瓜叶片碳代谢物质含量及酶活性 采用间苯二酚法测定蔗糖和果糖含量[13];高氯酸法测定淀粉含量[14];取黄瓜幼苗叶片0.2 g于液氮中速冻,采用上海杰美基因医药科技有限公司生产的试剂盒测定Rubisco酶活性[15];采用南京建成生物工程研究所研制的试剂盒测定蔗糖合成酶(SS)和蔗糖磷酸合成酶(SPS)活性。
1.2.3 黄瓜叶片和根系氮代谢物质含量及酶活性 硝态氮、氨态氮、可溶性蛋白含量测定参照文献[16-17],脯氨酸含量测定采用水合茚三酮比色法,硝酸还原酶(NR)测定利用磺胺比色法,谷氨酰胺合成酶(GS)测定参照文献[18],谷氨酸合成酶(GOGAT)活性测定参照文献[19]。每个处理重复3次。
1.3 数据分析
采用Excel 2003软件处理数据和绘图,采用DPS软件进行统计分析,并运用邓肯氏检验法进行差异显著性(P<0.05)检验。图表中数据均为“平均值±标准差”。
2 结果与分析
2.1 试验期间各处理夜间根区温度和气温的变化
由图2可知,试验期间温室内夜间气温总体呈上升趋势,3种处理根区温度受气温的影响也表现出升高趋势。5月26日试验处理结束时,适温处理、亚适温处理、低温处理的根区温度分别为19.8,14.6和9.8 ℃,较5月6日试验开始时的根区温度分别升高了1.5,1.2和1.7 ℃,但温度变化稳定在试验设定的范围内,符合试验对根区温度的要求。

图2 试验期间各处理夜间平均地温和气温变化Fig.2 Changes of root-zone temperature and air temperature during the experiment
2.2 根区温度对黄瓜幼苗叶片碳代谢及相关酶活性的影响
2.2.1 叶片淀粉、蔗糖及果糖含量 由表1可知,随着根区温度的降低,“黑籽”、“白籽”、“自根”的淀粉、蔗糖和果糖含量均表现出降低趋势,但嫁接苗的降低幅度小于自根苗。相同类型苗与其CK相比,低温处理下“自根”的淀粉、蔗糖和果糖含量降低幅度最大,分别较CK降低25.16%,33.88%和26.02%,而“黑籽”的淀粉、蔗糖和果糖含量降低幅度最小,分别较CK降低11.03%,26.82%和15.59%。适温处理与亚适温处理间“黑籽”淀粉含量差异不显著,亚适温和低温处理下“黑籽”、“白籽”、“自根”三者间的淀粉含量差异显著。3种温度处理下相同类型苗之间蔗糖、果糖含量差异显著;相同温度处理下“黑籽”与“白籽”、“自根”之间的蔗糖、果糖含量差异显著;低温处理下“白籽”与“自根”的蔗糖、果糖含量差异显著,其他温度处理下二者差异不显著。
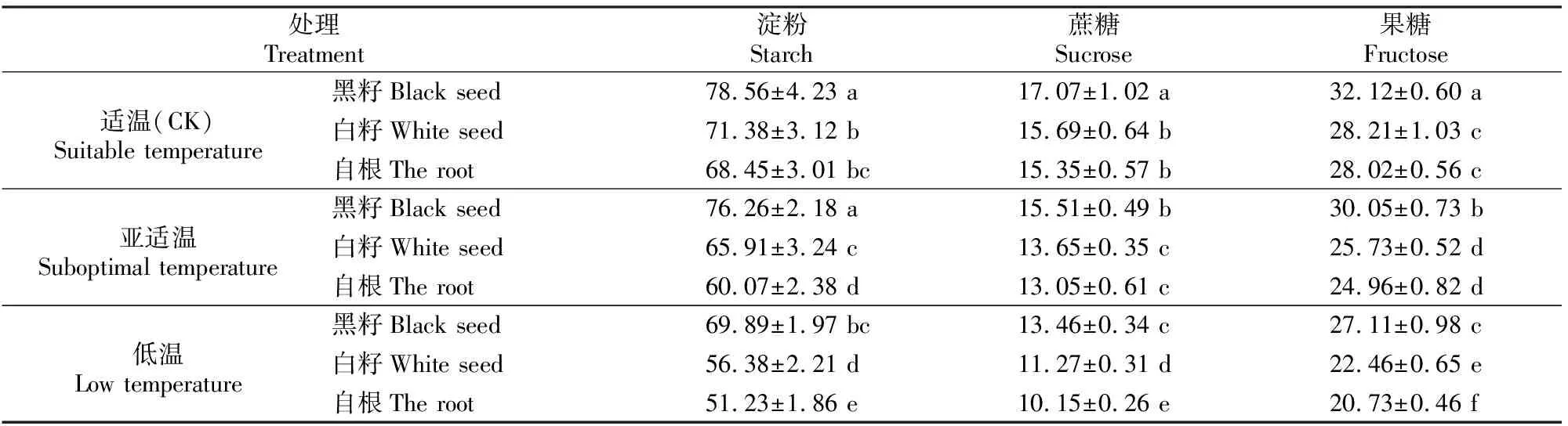
表1 根区温度对黄瓜叶片淀粉、蔗糖及果糖含量(干基)的影响
注:同列数据后标不同小写字母代表差异显著(P<0.05),下同。
Note:Different small letters in the same column mean significant difference atP<0.05 level,the same below.
2.2.2 叶片Rubisco酶、蔗糖磷酸合成酶(SPS)、蔗糖合成酶(SS)活性 根区温度对黄瓜幼苗叶片Rubisco酶、蔗糖磷酸合成酶(SPS)和蔗糖合成酶(SS)活性的影响如图3~5所示。图3~5显示,黄瓜幼苗叶片 Rubisco 酶、SPS、SS的活性均随处理温度的降低呈下降趋势。亚适温下,“黑籽”的Rubisco酶、SS与CK存在显著差异,分别较CK降低13.38%和26.49%;“白籽”和“自根”的Rubisco酶、SPS、SS活性与CK相比也存在显著差异,“白籽”的3种酶活性分别较CK降低20.66%,16.56%和29.24%,“自根”的3种酶活性分别较CK降低32.95%,22.37%和46.98%。低温处理下,“黑籽”、“白籽”和“自根”的Rubisco酶、SPS、SS活性与CK相比均存在显著差异,3种试材的Rubisco酶、SPS、SS活性从强到弱依次为“黑籽”>“白籽”>“自根”,“自根”的Rubisco酶、SPS、SS活性最低,分别较CK降低75%,55.24%和74.69%。
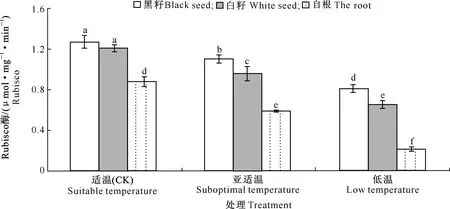
图3 根区温度对黄瓜幼苗叶片Rubisco酶活性的影响Fig.3 Effects of root zone temperature on Rubisco activities in cucumber leaves

图4 根区温度对黄瓜幼苗叶片蔗糖磷酸合成酶(SPS)活性的影响Fig.4 Effects of root zone temperature on SPS activities in cucumber leaves

图5 根区温度对黄瓜幼苗叶片蔗糖合成酶(SS)活性的影响Fig.5 Effects of root zone temperature on SS activities in cucumber leaves
2.3 根区温度对黄瓜幼苗叶片和根系氮代谢及相关酶活性的影响
2.3.1 叶片中硝态氮、铵态氮、可溶性蛋白和脯氨酸含量 根区温度对黄瓜叶片铵态氮、硝态氮、可溶性蛋白和脯氨酸含量(鲜基)的影响见表2。

表2 根区温度对黄瓜叶片铵态氮、硝态氮、可溶性蛋白和脯氨酸含量(鲜基)的影响Table 2 Effect of root zone temperature on contents (fresh basis) of soluble protein and proline in cucumber leaves
由表2 可知,亚适温和低温处理降低了黄瓜叶片中铵态氮、硝态氮含量,促使可溶性蛋白和脯氨酸含量显著升高。与CK相比,亚适温处理条件下“黑籽”的铵态氮和硝态氮含量未显著降低,“白籽”、“自根”的铵态氮和硝态氮含量显著下降,“白籽”分别较CK降低10.51%和6.62%,“自根”分别较CK降低15.74%和15.02%;亚适温处理条件下,“黑籽”、“白籽”、“自根”的可溶性蛋白和脯氨酸含量与CK相比也存在显著差异,其中“黑籽”的可溶性蛋白、脯氨酸含量最高,较CK分别提高15.51%和17.46%,“自根”的可溶性蛋白和脯氨酸含量最低,但也较CK分别增加11.91%和13.36%。低温处理条件下,3种试材的硝态氮和铵态氮含量较CK显著降低,脯氨酸和可溶性蛋白含量较CK显著升高,其中“黑籽”的铵态氮、硝态氮含量最高,分别较CK降低19.30%和13.28%,“自根”的可溶性蛋白和脯氨酸含量最低,分别较CK升高32.41%和36.11%。
2.3.2 根系中硝态氮、铵态氮、可溶性蛋白和脯氨酸含量 由表3可知,根区温度显著影响黄瓜根系中氮代谢相关物质的含量。与适温(CK)相比,亚适温处理显著影响“黑籽”和“白籽”的硝态氮、可溶性蛋白和脯氨酸含量,“黑籽”的硝态氮含量较CK降低17.12%,可溶性蛋白、脯氨酸含量较CK分别升高21.55%和22.82%;“白籽”硝态氮含量较CK减少17.72%,可溶性蛋白、脯氨酸含量较CK分别增加16.07%和20.16%;亚适温处理下,“自根”的硝态氮、铵态氮、脯氨酸和可溶性蛋白含量与CK相比也存在显著差异,铵态氮、硝态氮含量分别减少31.25%和22.22%,可溶性蛋白、脯氨酸含量分别增加14.90%和18.16%。低温处理下,与适温(CK)相比,3种试材的硝态氮、铵态氮、脯氨酸和可溶性蛋白含量均存在显著差异;“黑籽”的铵态氮和硝态氮含量显著高于“白籽”和“自根”,“自根”的铵态氮和硝态氮含量最低,分别较CK降低62.5%和46.33%;“自根”的可溶性蛋白和脯氨酸含量显著低于“白籽”和“黑籽”,“黑籽”的可溶性蛋白和脯氨酸含量最高,分别较CK提高56.03%和78.28%。
表3 根区温度对黄瓜根系铵态氮、硝态氮、可溶性蛋白和脯氨酸含量(鲜基)的影响Table 3 Effect of root zone temperature on contents (fresh basis) of soluble protein and proline in cucumber roots
2.3.3 叶片中硝酸还原酶(NR)、谷氨酰胺合成酶(GS)、谷氨酸合成酶(GOGAT)活性 由图6~8可知,与适温(CK)相比,亚适温和低温处理抑制了黄瓜幼苗叶片NR、GS和GOGAT的活性。亚适温条件下,“黑籽”的NR、GS和GOGAT活性与CK相比差异不显著,“白籽”和“自根”的NR、GS和GOGAT活性与CK相比均存在显著差异,“白籽”的NR、GS和GOGAT活性分别较CK降低9.13%,11.66%和6.25%,“自根”的NR、GS和GOGAT活性分别较CK降低15.77%,18.05%和7.55%。低温处理下,“黑籽”、“白籽”和“自根”的NR、GS和GOGAT活性与CK相比均存在显著差异,其中“黑籽”的NR、GS和GOGAT活性最高,“自根”的NR、GS和GOGAT活性最低。
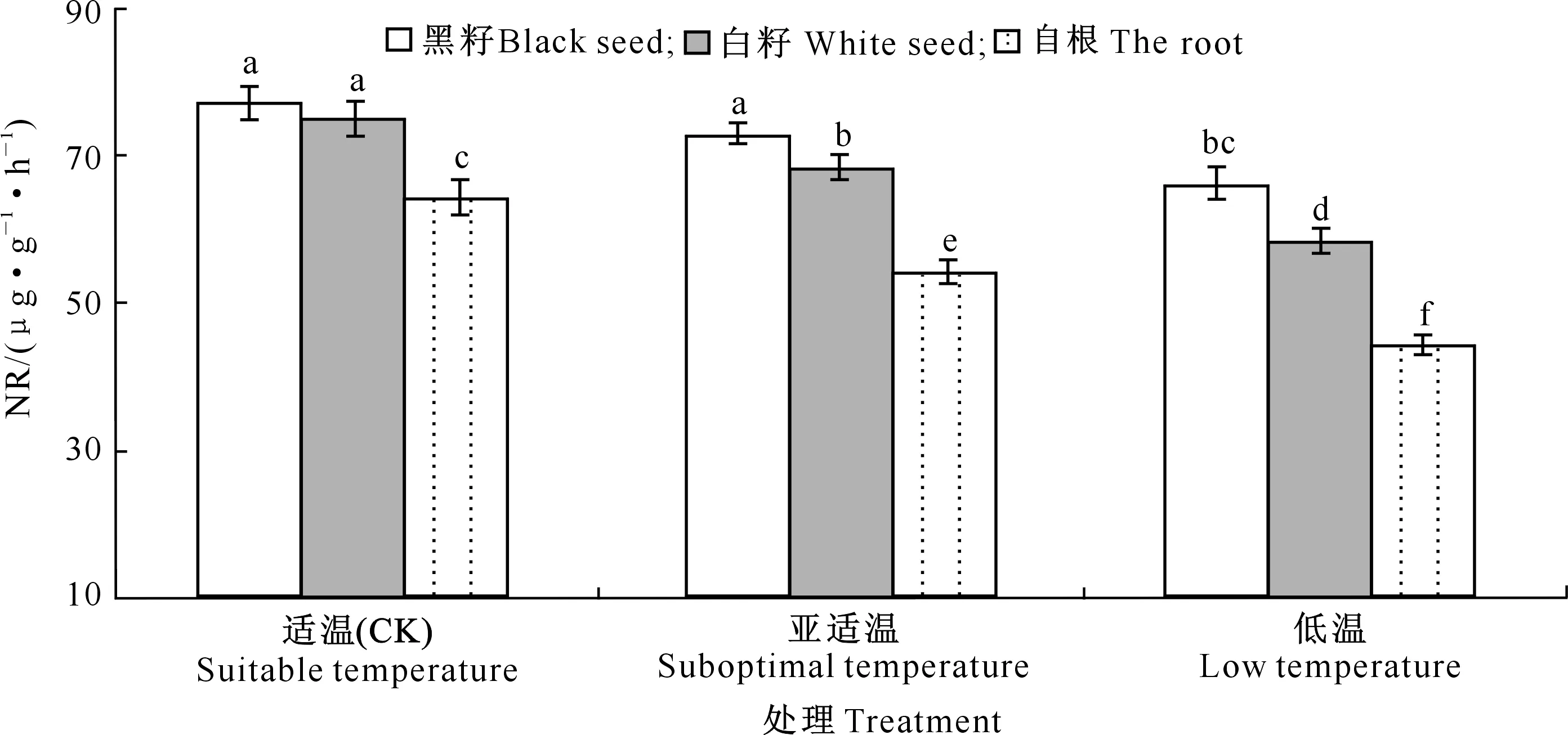
图6 根区温度对黄瓜幼苗叶片硝酸还原酶(NR)活性的影响Fig.6 Effects of root zone temperature on NR activities in cucumber leaves

图7 根区温度对黄瓜幼苗叶片谷氨酰胺合成酶(GS)活性的影响Fig.7 Effects of root zone temperature on GS activities in cucumber leaves

图8 根区温度对黄瓜幼苗叶片谷氨酸合成酶(GOGAT)活性的影响Fig.8 Effects of root zone temperature on GOGAT activities in cucumber leaves
2.3.4 根系中硝酸还原酶(NR)、谷氨酰胺合成酶(GS)、 谷氨酸合成酶(GOGAT)活性 由图9~11可知,黄瓜幼苗根系中NR、GS、GOGAT活性随着处理温度的降低逐渐降低。亚适温条件下,“黑籽”的NR和GS活性与CK不存在显著差异,但GOGAT活性与CK差异显著,较CK降低9.52%;“白籽”和“自根”的NR、GS、GOGAT活性与CK相比均存在显著差异,其中“白籽”的NR、GS、GOGAT活性分别降低16.31%,8.60%和19.04%,“自根”的NR、GS、GOGAT活性分别降低29.21%,10.28%和34.21%。低温处理下,“黑籽”、“白籽”和“自根”的NR、GS、GOGAT活性与CK均存在显著差异,“黑籽”的NR、GS、GOGAT活性最高,分别较CK降低18.17%,11.39%和25.39%,“自根”的NR、GS、GOGAT活性最低,分别较CK降低56.07%,24.57%和65.78%。

图9 根区温度对黄瓜幼苗根系硝酸还原酶(NR)活性的影响Fig.9 Effects of root zone temperature on NR activities in cucumber roots
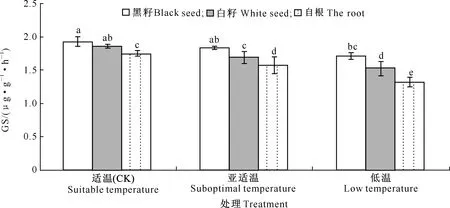
图10 根区温度对黄瓜幼苗根系谷氨酰胺合成酶(GS)活性的影响Fig.10 Effects of root zone temperature on GS activities in cucumber roots

图11 根区温度对黄瓜幼苗根系谷氨酸合成酶(GOGAT)活性的影响Fig.11 Effects of root zone temperature on GOGAT activities in cucumber roots
3 讨 论
黄瓜对根区温度非常敏感[20],不适宜的根区温度会破坏其地上部和地下部代谢平衡,由此会引发一系列的次生伤害。碳代谢在植物体的生命活动中起着重要的作用,其主要包括光合固定、碳水化合物的运输转化代谢以及碳积累代谢[21]。Rubisco酶活性是光合固定过程中的关键酶。蔗糖磷酸合成酶(SPS)和蔗糖合成酶(SS)则是糖转化与积累过程中所必需的酶[22]。研究发现,根区温度在20 ℃时,光合作用的相关酶活性高于13 ℃[23]。根区亚低温使黄瓜幼苗生长受抑制,磷酸烯醇式丙酮酸羧化酶、SPS和SS活性降低[10]。本试验中,与适温(对照)处理相比,亚适温和低温处理降低了Rubisco酶、SPS和SS 活性,这与前人的研究结果一致。本试验研究发现,亚适温和低温处理降低了黄瓜叶片淀粉、蔗糖及果糖的含量,淀粉含量的减少可能是由于根区低温抑制了黄瓜幼苗叶片SS和SPS活性,因此合成淀粉所需的底物腺苷二磷酸葡萄糖(ADPG)合成受阻,从而抑制了淀粉合成[24];而蔗糖和果糖含量的降低,其原因可能是根区低温导致黄瓜幼苗光合速率降低[25],同时SPS和SS活性下降,因此直接导致了光合产物的减少。

嫁接苗因为具有相对较强的抗逆境能力,所以已经被广泛应用在生产实践中。前人研究发现,砧木苗黑籽南瓜的生长和抗冷性优于新土佐[31],以黑籽南瓜为砧木的黄瓜嫁接苗,在低温下根系吸收养分与水分的能力显著强于自根苗[32]。本研究表明,相同根区温度处理下,“黑籽”碳氮代谢相关酶的活性及物质含量均高于“白籽”和“自根”;在亚适温和低温处理条件下,“黑籽”的可溶性蛋白和脯氨酸含量最高,“白籽”次之,“自根”最低。可溶性蛋白和脯氨酸可以有效缓解因低温胁迫导致的细胞内水分和膨压降低,从而维持细胞的正常功能,因此“黑籽”具有相对较强的抗低温胁迫能力。
4 结 论
亚适温和低温处理抑制了黄瓜幼苗碳氮代谢相关酶的活性,使得黄瓜幼苗体内淀粉、蔗糖和果糖含量减少,阻碍了铵态氮和硝态氮的吸收与利用。与此同时,低温处理诱导了植物体内可溶性蛋白及脯氨酸含量的升高,缓解了低温胁迫对黄瓜幼苗生理代谢造成的不利影响。“黑籽”受到根区低温胁迫的影响小于“白籽”和“自根”,其中“自根”受到的影响最大。
猜你喜欢
中老年保健(2022年3期)2022-11-21
农业现代化研究(2022年3期)2022-06-29
中国资源综合利用(2022年5期)2022-06-06
棉花学报(2020年3期)2020-08-08
科学与财富(2019年15期)2019-10-21
干旱地区农业研究(2017年6期)2018-01-31
中国生态农业学报(中英文)(2017年2期)2017-02-16
农家顾问(2016年5期)2016-05-14
植物营养与肥料学报(2014年4期)2014-04-08
中国蜂业(2013年18期)2013-01-17
