
珍稀植物青檀天然群体种子表型变异分析
2018-09-01 08:15唐国梁解孝满穆艳娟王丽丽
山东林业科技 2018年4期
唐国梁 ,解孝满 ,韩 彪 ,咸 洋 ,穆艳娟 ,陆 璐 ,王丽丽 ,武 香 ,董 昕*
(1.金乡县国有白洼林场,山东 金乡 272200;2.山东省林木种质资源中心,山东 济南 250102)
青檀(Pteroceltis tatarinowii)隶属榆科青檀属,是中国特有的单种属乔木树种,也是国家三级保护植物。青檀在我国东北、西北、华北和中南地区的19个省份均有分布,常生于山谷溪边的石灰岩山地,是石灰岩地区造林的先锋树种,也是优良的纸浆材,具有较高的生态价值和经济价值。长期以来,由于人们过于追求青檀的经济价值,使其天然林大量被采伐,许多遗传资源因此丢失。山东是青檀天然林受破坏较为严重的地区,对青檀天然群体进行保护迫在眉睫,目前青檀已被列为山东省重点保护的珍稀濒危树种之一。
搞清青檀群体的变异程度和遗传结构,对于保护和利用这一树种意义重大。表型多样性是遗传多样性研究内容中的一个重要部分,具有操作简单、快速和经济的特点,被认为是种群遗传多样性评价的有效手段之一[1]。天然群体的表型变异研究能够指示种群遗传变异的大小,是开展人工驯化及遗传育种工作的前提[2]。种子是林木进行繁衍的重要生殖器官之一,在选择压力下能够表现出很大的适应性[3]。种子形态作为较稳定的遗传特征,对于植物分类研究具有重要的价值[4]。目前,有关青檀的研究主要集中在群落结构特征[5,6]、繁育技术[7,8]、根际土壤[9]、木材纤维特征[10]、檀皮质量[11]、抗逆性[12]、分子标记分析[13]、诱变育种[14,15]、盆景制作[16]等,少有关于表型多样性的报道,本文对山东境内的5个青檀天然群体种子表型变异进行分析,揭示了山东青檀表型变异大小与变异规律,为其保护策略的制定、重要性状的遗传改良和良种选育提供理论基础。
1 材料与方法
1.1 试验材料
由于山东青檀天然资源遭到较为严重的破坏,现有青檀呈现少量集群分布和零散分布状态,林相较好的一般生长于保护区和寺庙内。2012-2017年,从山东主要山系陆续采种,采种群体位于北纬34°46′34″-36°38′42″, 东经 116°59′24″-117°32′33″之间,海拔高度108~350m。采集到5个群体33个单株的种子,采种母树胸径平均值大于25cm,种子来源情况详见表1。
1.2 表型性状的测量及分析
将果实阴干、去掉果翅,从每份种子中随机抽样30粒,用游标卡尺分别测量种子的横径、纵径、侧径,用量筒测量种子体积,用电子天平测量种子千粒重。其中,种子体积测量参照佘诚棋等[17]的方法,千粒重的测量采用百粒法,重复4次,所有测量结果均保留2位小数。
采用巢式线性模型[18]对群体间和群体内的表型差异进行方差分析。计算群体间方差分量与方差总量的比值,即表型分化系数,以此表示青檀群体间和群体内表型分化程度。表型分化系数计算公式为:。其中:VSI为表型分化系数;为群体间方差分量;为群体内方差分量[19]。利用SPSS 18.0和Microsoft Office Excel 2013等完成数据分析。
2 结果与分析
2.1 青檀种子性状的群体间变异
根据表2的方差分析结果可知,5个指示青檀种子形态的性状在群体间和群体内的差异均达到极显著水平。
从表3中可以看出,各群体的种子横径平均值为3.77mm~4.35mm,以莲台山群体的最大,佛峪群体的最小;各群体的种子纵径平均值为3.84mm~5.40mm,以曲阜群体的最大,灵岩寺群体的最小;种子侧径与种子横径和侧径相比变幅均较小,且以佛峪群体的最小;种子体积变幅也较小,以灵岩寺群体的平均值为最小,值为0.02cm3,各性状中以种子千粒重差异最大,平均值变幅为10.94g~23.12g,以曲阜群体的最大,灵岩寺群体的最小。
2.2 青檀种子性状的群体内变异
由表4可知,山东青檀种子性状的平均变异系数差异较大,各性状的平均变异系数为12.12%~47.00%。在5个表型性状中,种子体积的平均变异系数最大(47.00%),其次为种子千粒重(24.65%),种子横径的变异系数最小(12.12%),表明种子体积这一性状变异程度最大,而种子横径的变异程度最小,说明种子横径这一性状较其他性状更为稳定。此外,不同群体同一性状的变异系数存在差异,进一步对各群体的变异系数进行比较,各群体种子表型性状变异系数的顺序为:青檀寺 (25.86%)>佛峪(25.80%)>莲台山 (23.60%)>曲阜 (22.12%)>灵岩寺(16.31%)。高于平均变异系数(22.74%)的群体有青檀寺、佛峪和莲台山3个群体。其中,青檀寺群体的平均变异系数最大,说明该群体表型多样性最丰富;灵岩寺群体的平均变异系数最小,说明该群体的表型多样性最低。

表1 青檀天然群体采种母树来源

表2 青檀天然群体的种子表型性状方差分析表

表3 青檀天然群体种子表型性状的平均值和标准差
2.3 青檀种子性状的群体间表型分化
根据表5分析结果可知,群体间方差分量占总变异的27.37%,群体内方差分量占总变异的37.52%,机误的方差分量占总变异的35.11%;各性状表型分化系数的平均值为39.10%,表明青檀群体表型变异总量中有39.10%是群体间变异贡献的,另外的60.90%来自群体内变异,即群体内多样性大于群体间多样性。各性状的表型分化系数大小顺序依次为千粒重(59.70%)>种子纵径(58.97%)>种子横径 (42.79%)>种子体积 (26.14%)>种子侧径(7.90%)。
2.4 表型性状间的相关分析
由表6可知,各种子表型性状中大部分两两性状具有显著或极显著的相关关系,其中,种子千粒重和种子纵径与其他各性状均呈现极显著的正相关关系,种子横径与种子侧径呈极显著的正相关关系,种子纵径与种子体积呈显著的正相关关系。以上性状是能够表征青檀果实和种子的重要表型性状。

表4 青檀天然群体的种子表型性状变异系数表
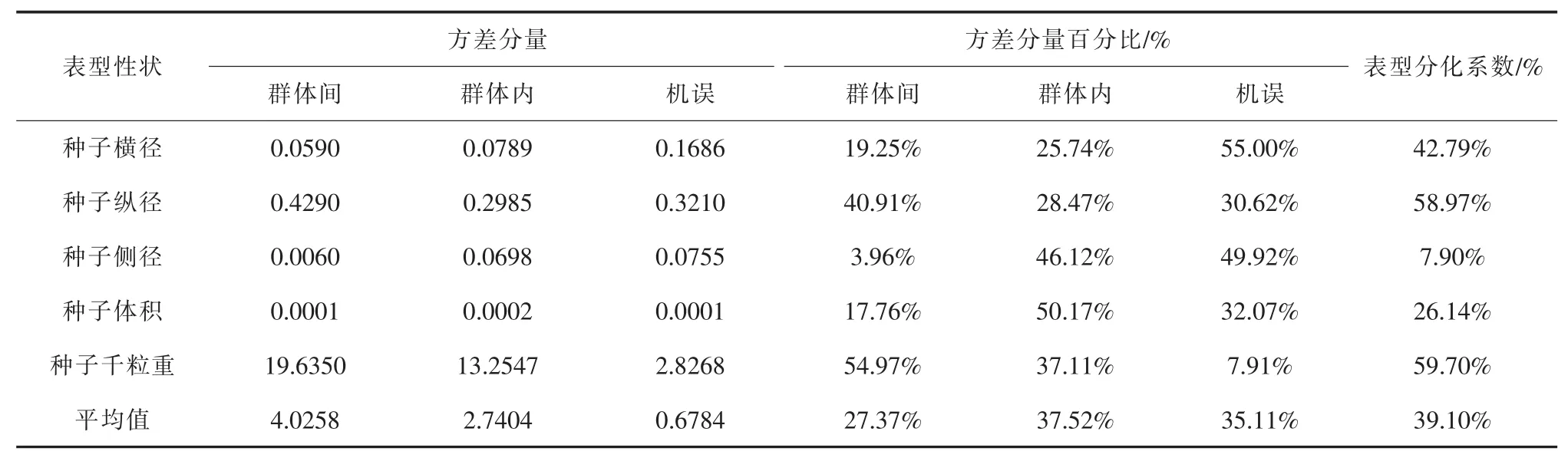
表5 青檀天然群体的种子表型性状方差分量及表型分化系数表

表6 青檀天然群体种子表型性状的相关分析和差异显著性检验
2.5 青檀群体表型聚类分析
采用欧氏平方距离系数进行聚类分析,5个青檀群体种子性状的表型数据自动聚合,生成聚类树系图,见图1。根据聚合水平不均匀发生跳变,可以将5个青檀群体明显划分成两组。莲台山群体和曲阜群体聚为一组,灵岩寺群体、青檀寺群体和佛峪群体聚为另一组。根据聚类结果有,地理上均是位于济南地区的莲台山群体没有优先与灵岩寺群体和佛峪群体聚类在同一个组,而是与距离较远的青檀寺群体优先聚合在一组,表明青檀群体没有依地理距离的远近进行聚类。综上,山东青檀天然群体的地理变异规律遵循随机变异模式。

图1 青檀天然群体表型性状聚类树系图
3 结论与讨论
研究表明,青檀的表型多样性非常丰富,各种子表型性状在群体间和群体内的差异均达到极显著水平。在各性状中,种子体积的变异系数最大为47.00%,种子横径的变异系数最小为12.12%。5个群体中,性状平均变异系数最大的是青檀寺群体,为25.86%,平均变异系数最小的是灵岩寺群体,为16.31%。说明青檀寺群体的表型多样性最丰富,灵岩寺群体的表型多样性最低,这可能是因为青檀寺群体受到破坏程度相对较小,其群体保存的相对完整,该群体的遗传基础较为广泛,在基因频繁交流的情况下,保持较高的变异水平。
山东青檀种子的表型变异中有39.10%来自群体间变异,有60.90%来自群体内变异,说明群体内多样性大于群体间多样性。尽管群体内的变异大于群体间的变异,然而,群体间的变异反映了地理、生殖隔离上的变异,群体间的多样性是种内多样性的重要组成部分[20]。5个青檀群体被聚合为两类,莲台山群体和曲阜群体聚为一组,灵岩寺群体、青檀寺群体和佛峪群体聚为另一组。地理距离相近的个别群体没有聚类在同一组,反而是与地理距离相差较远的群体聚类在同一组,说明青檀群体没有严格按照地理距离进行聚类,这可能是由群体生境的小地形差异导致的。
青檀是集生态、药用、饲用、材用等于一体的多种价值树种,蕴含巨大的开发潜力。长期以来青檀的生境受人为干扰严重,再加上被掠夺式的采伐利用,使其一度处于濒危状态。在山东省内已经很难找到大面积集中连片分布的天然青檀林,青檀的分布区域逐渐“岛屿化”、“片段化”[21],甚至在有些地区呈零散分布状态存在,保护野生青檀资源与保持青檀遗传多样性工作亟待进行。尽管本文是对青檀表型多样性的研究,然而表型是由遗传型与环境共同作用的结果,表型多样性丰富预示可能蕴含的遗传多样性丰富。该研究中青檀的表型变异十分丰富,且遗传变异主要存在于群体内。因此,对于青檀的保护可以采取原地保存策略,建立青檀原地保护区等,在保存青檀群体的同时要重视对群体内不同个体的保存。为进一步开发利用青檀的遗传变异,还要结合异地保存策略,尽可能多的 收集青檀全分布区内的种质材料,建立青檀种质资源库等。
猜你喜欢
中国防痨杂志(2022年6期)2022-11-25
江苏农业科学(2019年14期)2019-09-23
文萃报·周二版(2019年23期)2019-09-10
天津诗人(2018年4期)2018-11-14
商情(2018年5期)2018-03-28
现代园艺(2017年21期)2018-01-03
江苏农业科学(2017年7期)2017-05-23
文苑·经典美文(2016年8期)2016-05-14
中国康复理论与实践(2015年10期)2015-12-24
医学研究杂志(2015年5期)2015-06-10
