9种金线莲资源的染色体倍性及其核型分析△
2018-09-03 09:15谢卓宓牛欢古力王建明梁莹秦双双李胤嘉李明杰张重义
中国现代中药 2018年8期
谢卓宓,牛欢,古力,王建明,梁莹,秦双双,李胤嘉,李明杰,3*,张重义,3*
(1.福建农林大学 作物科学学院,福建 福州 350002;2.广西壮族自治区药用植物园,广西 南宁 530023;3.福建农林大学 作物遗传育种与综合利用省部共建教育部重点实验室,福建 福州 350002;4.福建农林大学 生命科学学院,福建 福州 350002)
金线莲Anoectochilusroxburghii(Wall.)Lindl属兰科开唇兰的一种多年生草本植物,别名金线兰、金丝草。金线莲作为我国传统的珍贵药材,具有除湿解毒、清热凉血、祛风利湿之功效,用于治疗咯血、膀胱炎、风湿性关节炎、急慢性肝炎、支气管炎、糖尿病等症[1-6]。金线莲在我国的分布主要于亚热带地区,福建省则是我国金线莲的标识性道地产区,由于金线莲种子萌发的苛刻性以及人工的过度采挖,导致现今的野生金线莲资源几乎处于濒危的边缘[7-10]。近几年,随着金线莲离体快繁技术的逐渐成熟,金线莲的人工种植面积正在逐渐扩大,栽培技术也相应逐渐成熟。但是在金线莲的栽培生产上,却存在种植面积广而分散、栽培品系多而不精、栽培技术散而不统等普遍问题。目前,金线莲品种选育较为杂乱,并且缺乏标准的品种选育体系,大部分栽培品种或品系为资源分离后直接转变而来,导致生产上金线莲同名异物、同物异名现象较为严重。同时,在金线莲的培育、生产和加工过程中,人们频繁使用来路不明的亲本,导致主要金线莲品种杂合退化现象较为严重。因此,详细的确定不同金线莲资源的遗传背景和遗传信息,彻底地排查不同地区金线莲资源亲缘关系和进化关系对于解决目前金线莲生产上的资源混乱的问题具有重要意义。现今,虽然在金线莲的研究中已经开发了诸如ISSR、SCAR、RAPD、SRAP等标记[11-13],分子标记方法在一定程度上为判明金线莲的不同资源间的亲缘关系提供了重要的分子手段。但这些标记体系由于其自身的缺陷,导致不能对现有金线莲资源体系,特别是亲缘关系较为相近的变异种而进行有效的区分,并且不能完全的反映出金线莲的遗传进化信息和背景信息。因此,为了更加精准的把握金线莲资源的遗传基础,准确的判明金线莲遗传进化规律,详细探明金线莲资源间倍性关系以及遗传差异将成为今后金线莲资源研究必要研究的方向之一。
不同物种细胞内染色体的组成和形态构成了该物种特有的细胞核型基础,呈现出了不同物种细胞的特定遗传形态,在进化的过程中不受外界环境因素干扰和影响。因此,一个物种或资源的核型模式在一定程度上反馈出了其进化的基本特征[14-16]。染色体核型分析是植物种质资源遗传学研究的重要内容,也是物种进化关系和系统分类的重要参考标准,在研究植物系统分类的起源、演化和鉴别中充当了重要的研究手段,目前,金线莲在细胞学上的研究相对较少,加之金线莲染色体形态小、数目多少变幅大等原因,使得金线莲核型分析较为困难,至今尚未有不同金线莲资源核型差异方面的研究内容。本文采用流式细胞仪方法对9个不同种质资源类型的金线莲品系倍性特征进行初步解析,同时,利用去壁低渗-火焰干燥法[17]对不同金线莲品系进行染色体制片和核型分析。本研究为进一步详细研究金线莲资源的类群划分和优良品系的选育提供重要的数据支撑和理论依据。同时,也为兰科植物的演化、进化特征和遗传规律的研究提供重要的细胞学佐证。
1 材料与方法
1.1 材料
选择福建等地区9种广泛种植的金线莲品种(系)资源作为实验材料,9种金线莲资源分别命名为GXHZ、GXJX、GXDY、FJQG、FJTN/1、FJTN/2、FJCY、FJSM、FJJY,所有金线莲种质资源材料均用组培体系进行保存,并由福建农林大学邹小兴(副教授)认证为金线莲(表1,图1)。


图1 9种不同种质资源金线莲表型图
1.2 方法
1.2.1 染色体条数的鉴定 采用去壁低渗-火焰干燥法[17]对不同品系金线莲的染色体数目进行鉴定分析。选取金线莲茎段接种后30~60 d的组培幼苗乳白色根尖,用涓流小心清洗掉组培苗根基上的残渣,放置于0.002 mol·L-18-羟基喹啉室温下避光处理2~4 h,之后转入现配的卡诺氏固定液(甲醇∶冰醋酸=3∶1)在室温下固定3~4 h。结束之后再用70%酒精清洗,并于-20 ℃保存。开始制片之前,将低温保存的材料恢复至室温后,滤去酒精,并用蒸馏水冲洗3次。截取根尖置于酶解液中,37 ℃恒温酶解1~1.5 h;加入冷蒸馏水冲洗后转入卡诺氏固定液中。用移液枪将截取的根尖均匀吸附到载玻片上,将玻片快速通过火焰,完成制片。将所制玻片经DAPI 染色后在荧光显微镜下进行观察,选择细胞内分散度相对较好的染色体,在100倍油镜下对分散较好的染色体细胞进行拍照。
1.2.2 核型分析及数据处理 以不同金线莲资源的细胞染色体拍摄图为基础,根据染色体长度、长短臂的差值、着丝点位置等进行同源染色体配对,并由长到短依序进行排序,绘制不同金线莲染色体的核型图。同时,根据单一细胞内不同染色体所测长度的平均值绘成核型模式图。根据李懋学等[18]的方法对不同资源金线莲的核型进行分析,观察同一资源材料根尖至少30个以上的细胞内的染色体条数,如果85%以上的细胞具有恒定一致的染色体个数,即可确证该资源材料的染色体具体条数。此外,参照韩春艳等[19]、Arano[20]的方法计算核型不对称系数,并利用Stebbins[21]的标准确定核型的类别。染色体分类(着丝点位置命名)则参照Levan[22]的命名法。具体计算公式如下:
(1)
(2)

(3)

(4)
(5)
(6)
1.2.3 染色体核型近似系数 根据谭远德、吴昌谋[23-24]等方法,选取染色体相对长度的极差、相对长度方差、臂比值的倒数均值、臂比值的倒数方差、着丝点指数均值等参数指标进行核型似近系数(resemblance-near coefficient,λ)和进化距离(evolution distance,De)计算,并利用Ntsys-pc2.1软件对不同金线莲资源进行聚类分析。
1.2.4 染色体倍性鉴定 以铁皮石斛DendrobiumofficinaleKimura et Migo标准二倍体为对照,取0.5 cm2金线莲新鲜待测植物叶片组织,置于培养皿中,同时,在植物叶片周围,加入400 μL Partec CyStain UV Precise P 裂解液,并用刀片将叶片切碎,以充分提取出完整的细胞核,提取时间设置为10 s。最后将切碎叶肉组织及裂解叶混合物用30 μm 滤网过进行滤,将过滤液收集至样品管内,并同步向样品管中加入Partec CyStain UV Precise P 染液1600 μL进行染色,时间为10 s。用CyFlow Space机型流式细胞仪(Sysmex Partec)结合上述提取液进行检测。
2 结果与分析
2.1 不同金线莲资源染色体的倍性分析
为了详细了解不同金线莲资源的遗传基础差异,本研究选取了9种叶片表型具有明显差异的金线莲资源(图1),通过流式细胞术对9个金线莲资源倍性进行初步判定。倍性分析时以遗传背景和倍性相对清晰的2倍体石斛材料作为对照,将对照的第一个荧光强度峰值位置设定在100(横坐标),随机选取每个金线莲资源组织中4000个细胞进行检测,根据细胞内第一个出现的荧光强度峰值与对照的荧光强度峰值的比值来判断其倍性(图2)。结果发现,9种不同金线莲种质资源在200~400(横坐标)位置之间的荧光强度皆有峰值,但所有材料的第一个荧光强度峰值位置基本处在200(横坐标),与对照材料比值为2,因此本研究所选用金线莲资源倍性初步判断皆为4倍体。

图2 9种不同金线莲流式细胞检测结果
2.2 不同金线莲资源的染色体形态分析
染色体条数和相应的核型组成是构成不同物种遗传信息基础,为了详细鉴定不同金线莲资源的染色体条数,本研究利用染色体核型分析技术详细鉴定了9种金线莲资源的核型组成。结果发现9种金线莲资源的染色体按照条数可以清晰的分为两组,一组为80条(2n=80),主要包括GXHZ 、GXJX 、FJTN/1、FJJY;另外一组染色体条数均为40条(2n=40),主要包括GXDY 、FJQG、FJSM、FJTN2、 FJCY(图3)。通过对不同金线莲资源染色体的形态进行仔细观察,发现9种金线莲资源材料染色体形态均较为微小,属于较为原始核型类型,着丝点位置不易分辨。其中,染色体中部着丝点(m)相对明显,未发现正中部着丝点(M)和随体。在GXHZ、GXDY、FJTN/1、FJTN/2、FJCY、FJSM、FJJY等金线莲资源中可以明显的观察到短染色体(S)、中短染色体(M1)、中长染色体(M2)、长染色体(L)等不同染色体类型;GXJX、FJQG等 2个金线莲资源中,观察到染色体大部分为中短染色体(M1)、中长染色体(M2)等染色体类型,没有观察到短染色体(S)(表2、图4)。
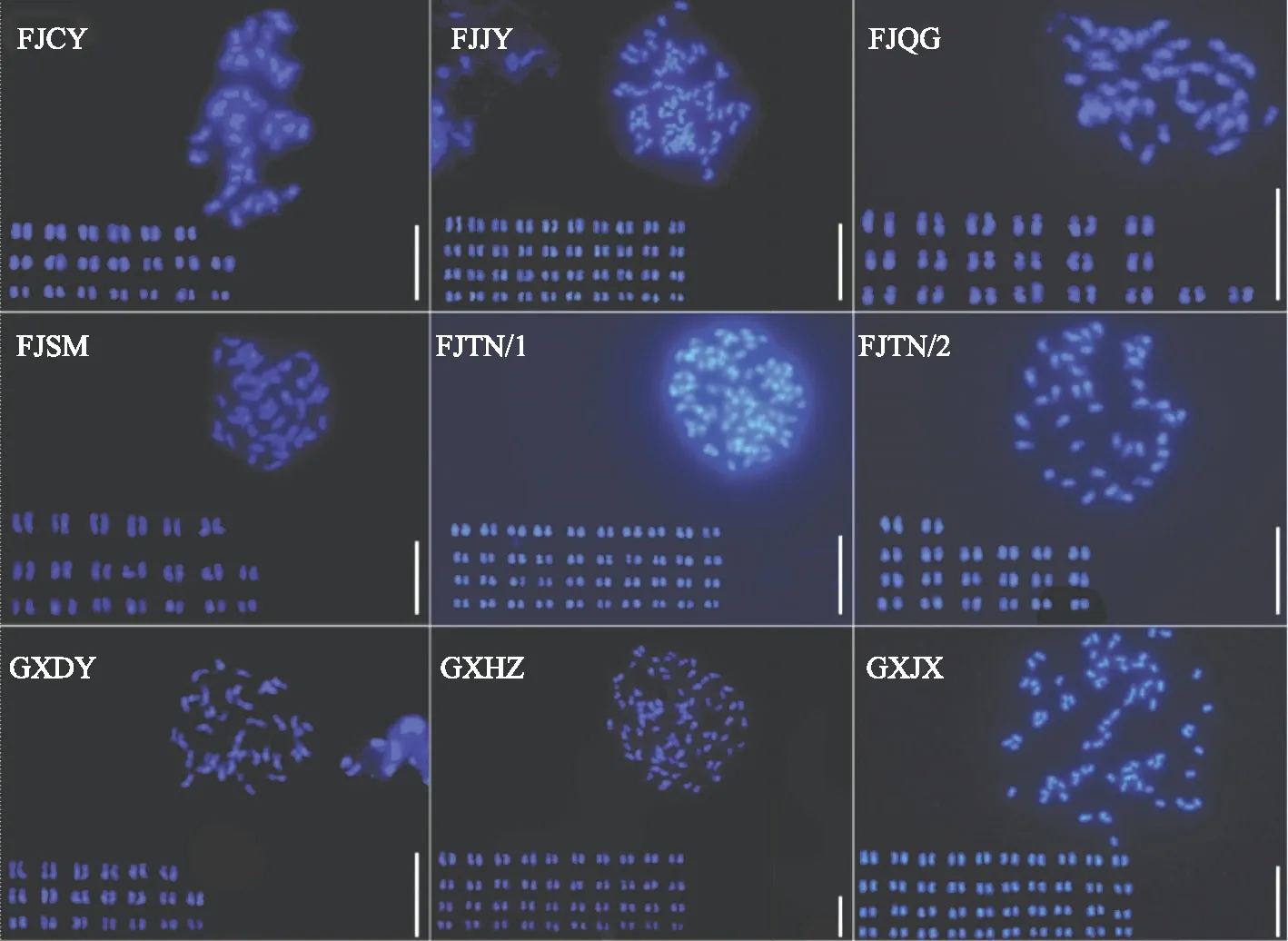
注:所有标尺为20 μm。图3 金线莲常规荧光形态及荧光核型
2.3 不同金线莲资源的染色体核型分析
在已获染色体核型的基础上,通过分析不同资源材料染色体长度、染色体长度比等不同参数可以初步的了解不同资源的遗传差异和进化秩序。同时,也可以根据具中部着丝点染色体的百分比与核型不对称系数的比值来分析物种的进化水平,比值越小表示样品进化水平越高。通过对9种金线莲的染色体长度分析发现,不同金线莲染色体长度变化不大。同时,分析臂比值大于2的染色体比例可以发现大部分材料比值位于0~0.325之间(表2)。臂比值可以衡量染色体对长度和对称性差异,可以从侧面反馈出物种进化进化程度。本研究9个金线莲资源臂比值大于2的染色体比例基本接近表明实验中所用材料进化程度较为接近。此外,通过分析不同遗传材料间染色体对称性和对称系数可以初步的解析不同遗传资源间的进化关系。本研究结合核型类型分析和核型不对称系数分析等方面的结果显示,核型不对称系数比较大的是GXDY、FJQG、FJTN/2与FJSM,数值都在60以上,其次的核型不对称系数也都在55左右,说明不同金线莲资源间遗传基础存在着一定的差异。此外,根据Stebbins等的核型分类法[21],不同植物染色体的核型按对称性程度的高低可分为 12种类型,即:1A,2A,3A,4A;1B,2B,3B,4B;1C,2C,3C,4C。从不同金线莲样品核型的参数进行综合判定分析,发现本研究9个样品绝大多数染色体的核型皆为2B型。
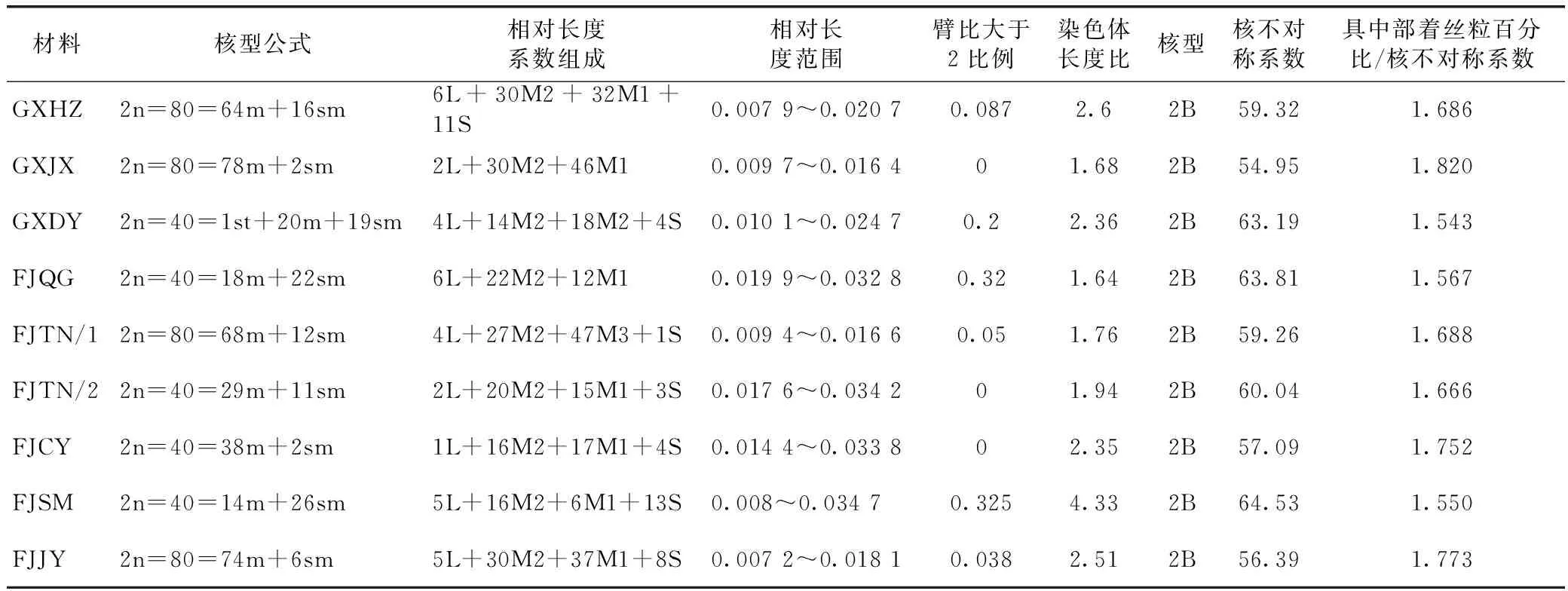
表2 金线莲染色体的核型参数

图4 金线莲核型模式图
根据核型似近系数的计算结果发现(表2,图5),9种金线莲资源可以明显的聚成3组,第一组主要包括FJQG、FJJY;第二组为FJTN/1、FJTN/2与GXJX;其余的为第三组。其中,FJJY与FJTN/2的似近系数数值最大为0.986,说明FJJY与FJTN/2的亲缘关系最近,或者可以说来源于同一资源后代。另外,FJSM与GXJX似近系数数值最小为0.473,表明 FJSM与GXJX亲缘关系最远。

表3 不同类型金线莲的核型似近系数(λ)和进化距离(De)
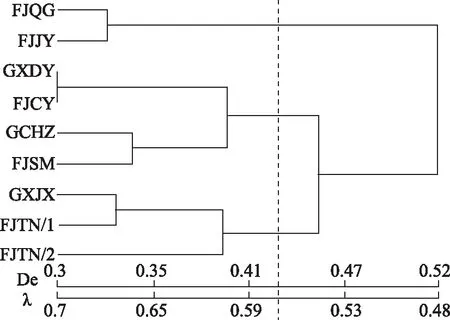
注:λ为核型似近系数;De为进化距离。图5 9种金线莲核型似近系数聚类分析
3 讨论
金线莲属于典型兰科植物,有着较为丰富的遗传资源,但是现今研究主要集中在金线莲药效成分分析、离体快繁、栽培技术方面[25-27],有关于金线莲染色体鉴定及核型分析等相关方面的研究较少。曾雅娟[28]利用常规压片的方法发现金线莲染色体数为2n=40,核型公式为 2n=40=38m+2sm,属于2A型核型。王丽芳等[29]用同样的方法鉴定金线莲的染色体数为40条,并用秋水仙素将植株染色体数成功加倍为80条。本研究选择了福建地区分布较为广泛、代表性较强、性状差异较为明显的9种金线莲资源,从细胞学水平上详细鉴定不同金线莲遗传资源间染色体条数及其核型构成差异。结果发现9种金线莲资源的染色体条数为2n=40和2n=80条2种类型。其中染色体数目为2n=80的类型主要包括GXHZ、FJTN/1、FJJY和GXJX 4个金线莲资源;染色体数目为2n=40的类型主要包括FJCY、FJQG、FJSM等5个金线莲资源。通过细胞倍性分析发现,2个 类型金线莲资源皆为4倍体,流式细胞术分析结果同样证实了所选金线莲资源均为4倍体。在同一地区金线莲资源会出现染色体数量的多寡变化可能受到多种因素的影响,比如:可能是由于金线莲在进化过程中的自然加倍或自然杂交造成的;也可能是金线莲在人工选育时人工培育体系造成的;也可能是因为不同的物种或是同一个物种的不同细胞之间有丝分裂的各个时期不同而产生变化。此外,目前大部分金线莲品种选育缺乏系统的育种程序,大部分品系均来自于野生资源的优良变异类型或野生材料的多代培育材料。不同研究人员在采样或繁殖时,通常会采用花粉直接繁殖用于快速保存所获的野生资源,这种方式也可能会造成倍数型差异。金线莲倍性差异为丰富金线莲的遗传资源和培育优异金线莲品种提供了重要的亲本材料。本研究所用金线莲种质资源大多采自林下的野生生态环境,核型构成存着较大差异,这种差异反馈出金线莲遗传资源的多样性和进化的特殊性。
在被子植物中核型进化往往是由对称向不对称的方向变化,在系统演化位置上越古老的物种其核型越对称,越进化的物种,其核型越偏向不对称[26]。根据李懋学[18]的核型分类标准,本研究所获取的9种金线莲资源均为2B型,与曾雅娟[28]等所鉴定的2A型不同,这可能主要是由于在研究金线莲的核型构成时所选取的研究材料不同所致。在金线莲核型分析中,由染色体平均长度可知,金线莲染色体属于小染色体类型,并且未发现随体的存在。在物种核型研究中,可以用具有中部着丝点染色体的百分比与核型不对称系数的比值来衡量物种的进化程度,比值越小也就越进化[30]。本研究所鉴定的9个金线莲资源的核不对称系数大部分位于54~65之间,进化起源上相对较早。9种不同种质资源在进化程度上各有差异,而这些差异有可能是因为他们的染色体组型特征或者血缘组成的差异导致。同时,这种差异也有可能在不同程度上反应出了不同种质资源类型在生态适应性上的差异,比如:FJQG、FJSM进化程度较为先进,其生态适应性可能更好,可能更适合于引种。通过对样品的核型分析,我们可以对今后的金线莲引种提供了一项可靠的依据。而且,物种的进化是以适应生境的变化为前提的,外界生存环境的变化是物种进化的直接驱动力。因此,结合物种进化的先进程度,可以从侧面窥视出物种随着自然环境变化与其自身在遗传水平上的响应、调整和适应的变化规律。根据物种的这种规律尺度反推重现则可以为药用植物仿野生栽培技术体系中品种的选择和契合生境的优化提供重要的参考依据。
目前,在药用植物栽培中,详细的追寻栽培药用植物进化的生态足迹,了解野生药用植物到栽培药用植物之间生境变迁规律,实行仿野生栽培技术体系的构建,已经成为实现药用植物的药性回归和安全生产的重要技术保障。金线莲大多数栖息于林下阴生环境,决定其进化耐阴性和对林下复杂生态环境的适应性。特别是随着金线莲仿野生栽培技术的逐渐推广,如何筛选和培育出耐阴抗逆性较强的种质资源已经成为金线莲发展的重要瓶颈性问题。因此,后续的研究可以将植物形态学特征、细胞遗传学依据与现代分子生物学相结合,将会更加准确地对金线莲资源的系统进化以及血缘关系作出更加客观的评价,为培育优质、适产和高适应的金线莲品种奠定基础。
致谢:感谢作物生态与分子生理学福建省高校重点实验室(福建农林大学)对本研究支持;感谢福建农林大学王凯教授课题组对金线莲染色体核型研究的指导。
猜你喜欢
小学生作文(低年级适用)(2022年11期)2022-12-02
云南医药(2021年6期)2022-01-08
临床检验杂志(2021年10期)2021-11-24
中国现代中药(2021年8期)2021-10-09
川北医学院学报(2021年6期)2021-07-13
科学之谜(2019年3期)2019-03-28
科学之谜(2018年8期)2018-09-29
中成药(2017年7期)2017-11-22
源流(2017年9期)2017-10-29
恋爱婚姻家庭·养生版(2016年9期)2016-09-07