
一个新的小麦-中间偃麦草的部分双二倍体及其白粉病与条锈病的抗性鉴定研究
2018-09-04 08:38聂林曼付体华
四川农业大学学报 2018年3期
杨 园,聂林曼,付体华
(四川农业大学农学院,成都 611130)
普通小麦(Triticum aestivum L.)的改良依赖于丰富的遗传多样性,而小麦育种中所需的大多数有利遗传变异都能够由小麦族的野生近缘种作为基因源提供。远缘杂交是一种有效的植物物种间的基因转移手段,在基因由小麦野生近缘种向普通小麦转移的过程中尤为重要[1-2]。利用小麦与其野生近缘种的远缘杂交,育种者已合成了大量双二倍体或部分双二倍体。这些双二倍体是重要基因转移的中间载体,可以用于从小麦野生近缘物种向普通小麦中导入优良性状基因[3-4]。中间偃麦草(Thinopyrum intermedium,2n=6x=42,JJsSt)是一种多年生的异源六倍体草本植物,它具有许多优良的遗传多样性,是小麦遗传改良中一个重要的三级基因源[5-6]。过去的几十年中,许多的属间杂种和细胞遗传系被创制并用于小麦育种中,这些品系包括不同小麦与偃麦草的部分双二倍体、外源易位系、代换系以及附加系[4,7-12]。基因组原位杂交(GISH)技术具有简单、准确以及高效的特点,被广泛地运用于分析小麦与小麦野生近缘种的杂种后代的遗传多样性[13]。基于拟鹅观草DNA(St染色体组)作为探针的GISH技术可将中间偃麦草染色体按杂交信号分布划分为J,Js和St 3个染色体组[14]。中间偃麦草是一种自然杂交形成的多年生物种,据研究不同来源的中间偃麦草其染色体组成也不尽相同[15-17]。现有的小麦-中间偃麦草部分双二倍体并没有包含中间偃麦草所有染色体[3]。因此,为了将中间偃麦草的优良基因整合到小麦中,我们仍需要不断创制和选育新的小麦-中间偃麦草部分双二倍体。到目前为止,已经有一些双二倍体被培育出来,并在分子细胞遗传学层次上对其染色体组结构进行了鉴定,如TAF46、中字系列部分双二倍体、Otrastsyuskaya(OT)、78829、TE-3、E990256、TE253和TAI8335等,这些双二倍体所拥有的中间偃麦草染色体都存在一定的差异[3,6,18-21],其中大多数已经被广泛地应用于小麦改良中[5,22]。总的来说,这些被报道的部分双二倍体具有旺盛的生长能力和高结实率,多数拥有56条染色体,通常由完整的小麦染色体组附加一个杂合的外源染色体组构成[3]。也就是说包含56条染色体的小麦-中间偃麦草部分双二倍体其外源染色体数目都不少于14条。品系12-1179是我们实验室近年来选育的一个小麦-中间偃麦草部分双二倍体,不仅对当前我国流行的条锈病和白粉病具有高度的抗性,而且其包含的外源染色体数目少于14条,这表明它是一个携带不同染色体结构的新的部分双二倍体。在本研究中,我们希望通过GISH和FISH技术确认其染色体结构和外源染色体来源,并对其潜在的白粉病和条锈病抗性进行评价,为其在遗传育种研究的应用奠定基础。
1 材料和方法
1.1 材料
供试材料包括拟鹅观草品系PI 232131(Pseudoroegneria spicata,St染色体组,2n=2x=24),小麦品种中国春、绵阳26、品系SY95-71和小麦-中间偃麦草双二倍体品系12-1179,其中品系12-1179由四川农业大学农学院细胞遗传实验室创制、筛选和保存。该品系是以中国春小麦作为母本与中间偃麦草杂交得到F1代杂种,再与小麦栽培种绵阳26回交得到F2代杂种,从F2代杂种自交筛选结实性好的植株繁殖到F8代。拟鹅观草种子来源于美国国家小粒作物收集中心。品系SY95-71来源于四川农业大学小麦研究所的小麦,具有高度感染条锈病和白粉病能力[23],作为感病对照。
1.2 方法
1.2.1 细胞学鉴定
待种子在培养皿中生长到根尖约1.5~2 cm时,将根尖剪下放入一氧化二氮(N2O)中处理2 h,然后用90%的冰乙酸固定10 min,将其储存于70%的乙醇中并于4℃冰箱中保存待用[24]。有丝分裂染色体制片根据Han F.等[25]的方法稍加改动制备。减数分裂制片是采取处于减数分裂中期Ⅰ的麦穗并将其固定在卡洛固定液Ⅰ中,用品红对花粉母细胞进行染色。GISH采用拟鹅观草DNA作为探针,而FISH采用Oligo-pSc119.2-1和Oligo-pTa535-1作为探针[26]。拟鹅观草DNA采用地高辛(digoxigenin-11-dUTP)标记,具体步骤按照地高辛说明书操作,中国春小麦DNA作为封阻。GISH的操作步骤根据Chen Q.等[14]和Han F.等[27]的描述进行。寡核苷酸探针Oligo-pSc119.2-1和Oligo-pTa535-1的5'端分别用6-carboxyfluorescein(6-FAM)和6-carboxytetramethylrhodamine(Tamra)进行标记[26],由上海 Invitrogen Biotechnology公司合成。合成的寡核苷酸探针用1×TE缓冲液溶解和稀释,稀释倍数按照Tang Z.X.等[26]的描述进行,FISH程序按照Han F.等[25]的描述进行。利用带有CMOS相机以及Spotsoftware(V.4.6)的荧光显微镜(Nikon Eclips80i)进行照相。
1.2.2 抗病性鉴定
品系12-1179以及其亲本的白粉病与条锈病抗性测试分别在四川农业大学雅安农场进行。小麦品系SY95-71属于高感条锈病与白粉病的感染型(infection types,ITs),在本实验中被用作阳性对照评价小麦感病程度。通过在苗期接种3个白粉病菌株(E20,E21和race 15)来评价测试小麦品系的白粉病抗性,而在成株上接种当前流行的混合的条锈病菌(包括条中 30,31,32及水源-46)评价测试小麦品系的条锈病抗性,条锈病菌由四川农业科学院植物保护研究所提供。当SY95-71高度感病时,按照0到4级评价体系记载所鉴定品系的白粉病和条锈病抗性[28-29]。
2 结果与分析
2.1 品系12-1179的形态学以及农艺性状调查
成株12-1179表现为小麦-中间偃麦草中间形态,但更加偏向于其小麦亲本(图1a)。它具有直立生长的习性,成株高度在95~105 cm,矮于中国春小麦(125 cm)而高于绵阳26小麦(88 cm)。在田间,每个植株平均能产生4~6个有效分蘖;穗长为14~17 cm,每个麦穗具有17~20个小穗数,小穗在穗轴上分布较稀疏(图 1b),平均结实率70%左右。种子的形态像中间偃麦草,种子细长但比较饱满(图1c),千粒重为28 g左右,界于小麦与中间偃麦草之间的水平。

图1 小麦栽培种绵阳26与品系12-1179的形态特征Figure1 The morphology features of wheat cultivar Mianyang 26 and amphiploid line 12-1179
2.2 品系12-1179的细胞学鉴定
对28株12-1179材料进行了根尖染色体计数。不同植株的染色体数目为54~57条,78.57%的植株染色体数目为56条(表1)。在部分非整倍体(2n=54或55)中可观察到1~2个端着丝粒染色体(表1)。对12-1179植株(2n=56)减数分裂中期Ⅰ染色体的配对行为进行分析发现有54.05%的花粉母细胞(PMCs)能形成28个二价体(图 2a),而在其他花粉母细胞中单价体为0~8个。少量的花粉母细胞中偶尔会观察到一个三价体或四价体(图2b)。通过对175个花粉母细胞(2n=56)减数分裂的统计,其减数分裂配对构型为1.33Ⅰ+27.26Ⅱ+0.01Ⅲ+0.03Ⅳ(表2),表明品系12-1179在减数分裂传代过程中基本保持稳定。
使用拟鹅观草DNA为探针,中国春小麦DNA为封阻的GISH技术对品系12-1179的外源染色体进行了鉴定,28株12-1179材料的外源染色体组成结构列于表1。结果表明所有12-1179植株的根尖细胞都仅含有12条中间偃麦草染色体(图3a),未发现小麦-中间偃麦草易位染色体。根据GISH信号分布特点[14],可将这12条外源染色体区分成中间偃麦草不同的染色体组。其中,6条染色体上都有强烈的杂交信号,属于St染色体组,4条染色体着丝粒和端粒有强烈的杂交信号,属于Js染色体组,而余下2条染色体一条臂在端粒处有杂交信号,而另一条整臂和着丝粒处都有杂交信号,属于St-Js易位染色体(图3a),易位染色体的易位点需进一步研究确定。在12-1179中没有发现J染色体。对减数分裂染色体的GISH分析表明减数分裂中期Ⅰ的花粉母细胞有6个二价体具有杂交信号(图3b)。说明品系12-1179的12条外源染色体能够在减数分裂期间正常配对,表明12-1179的外源染色体由3对St,2对Js以及1对St-Js易位染色体构成。
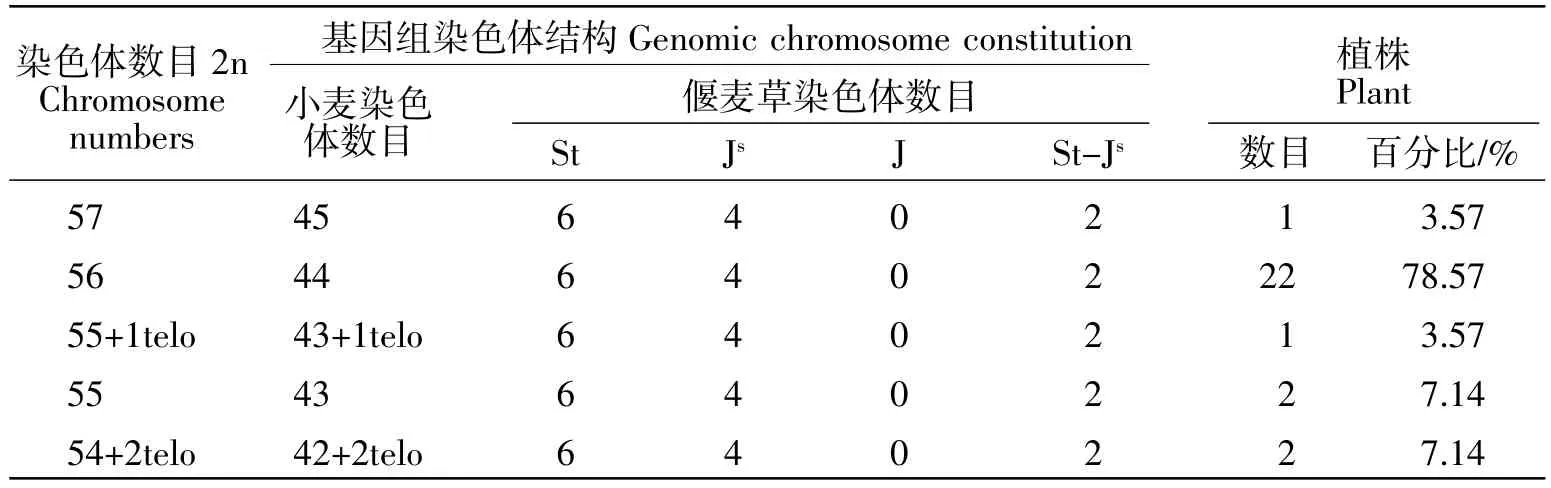
表1 28个12-1179植株的染色体数目和结构Table1 Chromosome number and constitution in 28 plants of line 12-1179
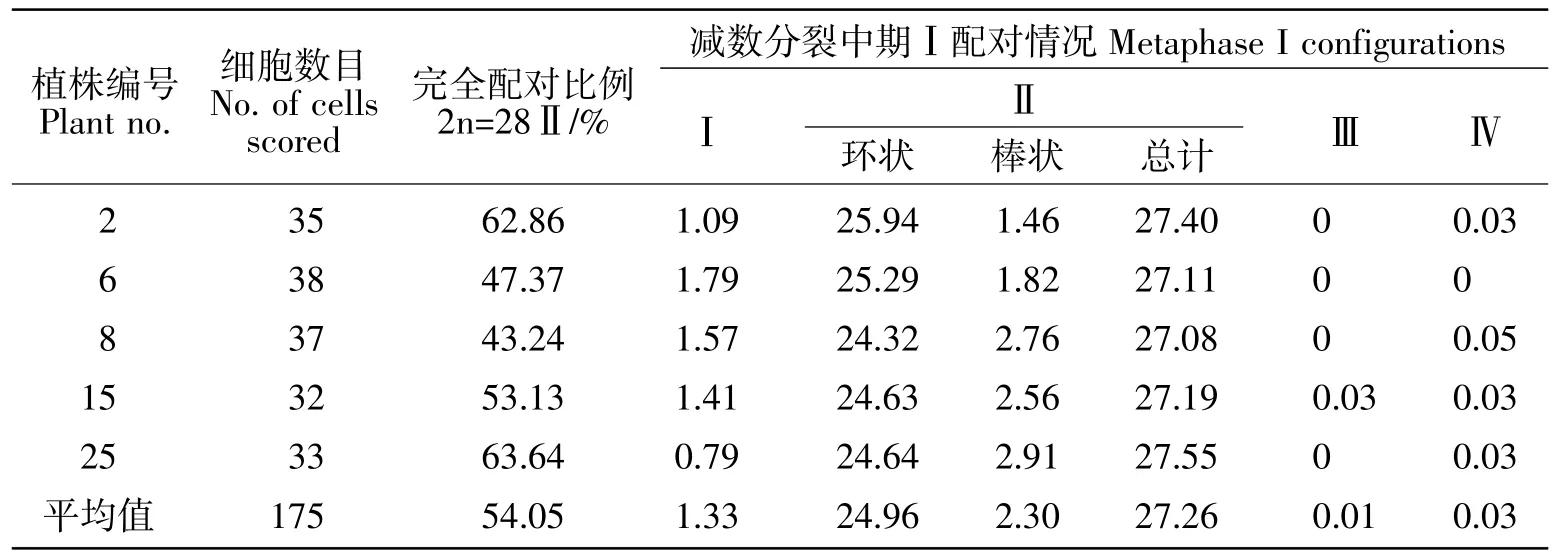
表2 5个12-1179(2n=56)植株减数分裂中期I的配对统计Table2 Chromosome pairing at metaphase I in five 56-chromosome plants of line 12-1179

图2 品系12-1179的减数分裂中期Ⅰ的结构Figure2 Meiotic configurations at metaphaseⅠof line 12-1179
使用寡核苷酸探针Oligo-pSc119.2-1和OligopTa535-1对品系12-1179进行FISH分析以观察小麦染色体数目及组成。结果表明12-1179具有完整的A和B染色体组,而在不同12-1179植株中有不同数目的D染色体。根据统计,在12-1179植株(2n=54-57)中,5D 染色体数目为 2~5条(图 3c~图3e),而在2n=56的12-1179植株中,5D染色体数目为4条(图 3c)。此外,在非整倍体(2n=54-55)植株中观察到的端着丝粒染色体都为5DL(表1和图3e)。因此12-1179中缺失的一对中间偃麦草染色体应属于第五同源群,其功能在很大程度上由增加的2条5D染色体补偿。所以品系12-1179的外源染色体组的组成是6St+4Js+2Js-St+2*5D。根据已有报道,小麦-中间偃麦草部分双二倍体的外源染色体组一般只由中间偃麦草的3个染色体组构成[3],因此,品系12-1179属于一个新的小麦-中间偃麦草部分双二倍体。
2.3 抗病性鉴定结果
利用当前流行的白粉病菌株和混合的条锈病菌对品系12-1179及其亲本进行了抗病性鉴定。从田间表现来看,中间偃麦草与12-1179对所有接种的白粉病与条锈病都免疫,而小麦栽培种中国春和绵阳26都表现高感(表3)。由于品系12-1179的小麦亲本都高感白粉病与条锈病,因此,12-1179的白粉病和条锈病抗性很可能来源于它的供体亲本-中间偃麦草。
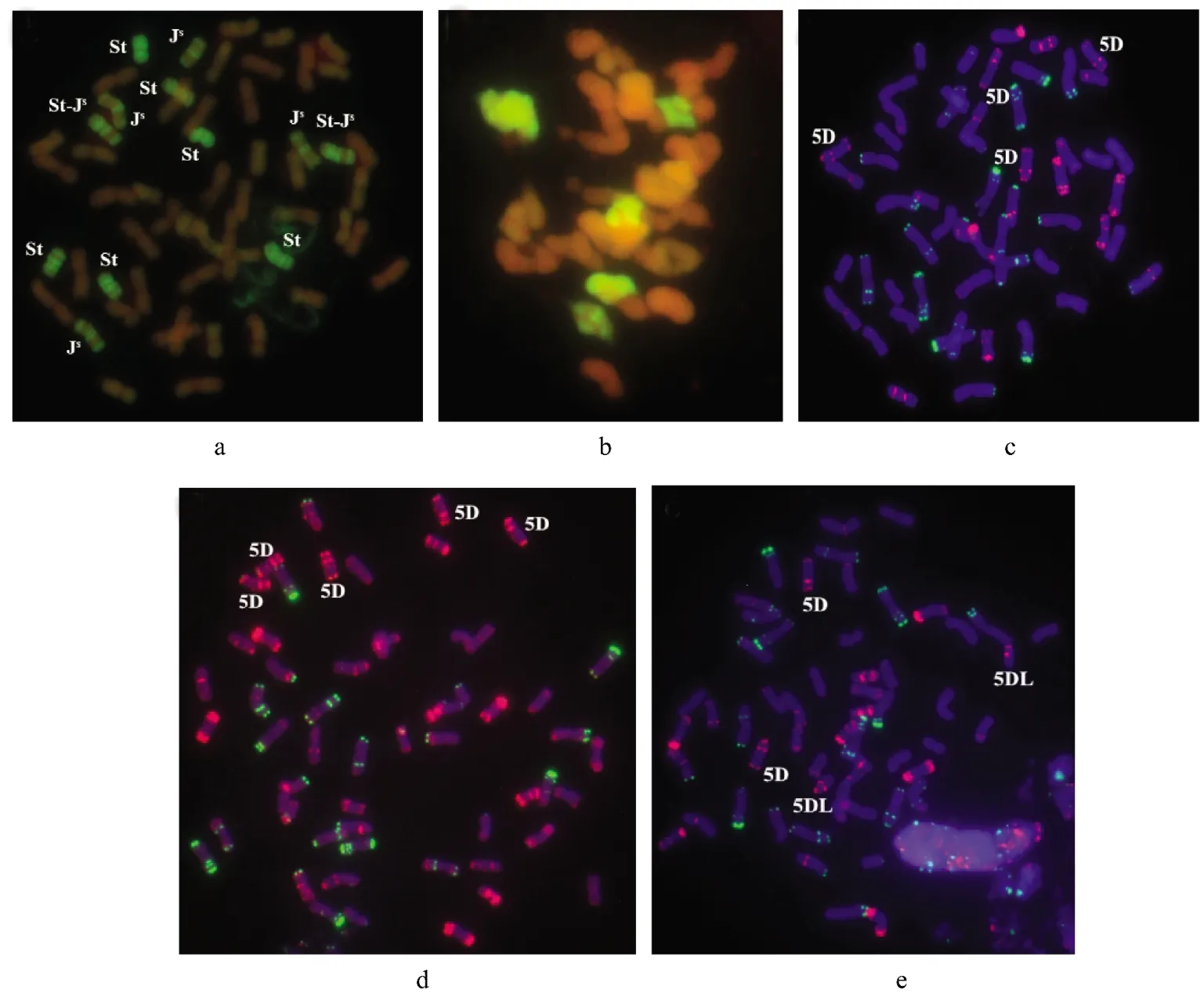
图3 品系12-1179植株的GISH和FISH分析Figure3 GISH and FISH patterns of line 12-1179
3 讨论与结论
对合成的小麦-外缘物种双二倍体进行细胞学的稳定性评价在育种过程中具有理论指导和实际意义。一般而言合成的小麦-中间偃麦草双二倍体都具有较稳定的减数分裂行为,其二价体频率高而多价体频率低,同时具有良好的结实率[18]。G.Fedak等[3]报道不同的小麦-中间偃麦草部分双二倍体减数分裂中期I的二价体频率变化在25.12~27.46之间,而单价体频率在0.43~2.55之间。在我们所研究的2n=56植株中,由于存在4条5D染色体,理论上每一个减数分裂细胞中至少应发现一个四价体,然而根据175个花粉母细胞(2n=56)统计分析发现54.05%的细胞能形成28个二价体,平均每个细胞有1.33个单价体,而每个细胞平均出现0.01个三价体和0.03个四价体,其多价体的数量远小于预期(表2)。表明品系12-1179减数分裂中期Ⅰ的染色体配对行为与已报道稳定的部分双二倍体相似[6,13,18],也类似于稳定的普通小麦缺体-四体的配对行为[30]。这表明品系12-1179在细胞学上十分稳定,因此具有比较高的结实率,这些结果说明品系12-1179的染色体遗传组成在很大程度上是平衡的[3]。

表3 12-1179和其亲本的条锈病与白粉病的抗性鉴定Table3 Reactions to stripe rust and powdery mildew in partial amphiploid12-1179 and its parents
了解部分双二倍体的外源染色体对于高效地将其优良性状转移到小麦中进行改良是非常重要的。因此,不同学者采用不同手段对选育的小麦-中间偃麦草部分双二倍体染色体组成进行鉴定。根据报道,所有小麦-中间偃麦草双二倍体(2n=56)的外源染色体都不少于14条,其中至少包括St,J和Js中的两种,且小麦染色体至多42条[3,21]。在本研究中,通过GISH分析表明品系12-1179的外源染色体只有12条,其中包括3对St,2对Js和1对Js-St易位染色体。品系12-1179的外源染色体少于已有报道双二倍体(2n=56)的外源染色体数目(图 3a)。FISH分析表明,12-1179包含4条5D染色体(图 3c),但能进行稳定地减数分裂且结实率达70%。由此我们推测中间偃麦草染色体组中的一对外源染色体由两条5D替换,而且外源染色体缺失的功能由5D染色体补偿,因此这个复合的外源染色体组不仅包含中间偃麦草染色体,还应该包含两条5D染色体。12-1179具有不同于已报道的部分双二倍体,如中字系列[19],TAF46[31],78829[32],OT[18],TAI8335[6]和 TE253[13]等的染色体组成,以上部分双二倍体都拥有一个由14条不同中间偃麦草染色体组成的复合染色体组,因此12-1179是一个新的小麦-中间偃麦草部分双二倍体。
在农艺性状方面,成株12-1179与小麦亲本更相似,却没有旺盛的生长习性和分蘖能力,小穗数也偏少,可能原因是第五同源群的中间偃麦草染色体被小麦5D替换所致。我们知道调节小麦春化作用以及区分小麦为春性、冬性和半冬性基因Vrn位于小麦第5同源群染色体长臂上,特别是5AL和5DL[33-35]。Vrn基因负责调控小麦营养生长和生殖生长之间的转换[36]。中国春小麦5D缺体是唯一导致小麦产生冬小麦习性且使抽穗期延迟的缺体小麦[37],所以小麦染色体5D替换了中间偃麦草的第五同源群染色体,导致品系12-1179的冬性特征减弱而更类似于春小麦亲本且分蘖能力较弱。
中间偃麦草作为一种重要的多年生小麦族物种,对多种真菌病害有良好的抗性以及对不利环境有很好的耐受性[16,19,38]。因此,中间偃麦草在小麦遗传改良中能作为抗性基因的供体被广泛地使用。迄今为止,抗小麦条纹花叶病毒(WSMV)[39]、抗大麦黄矮病毒(BYDV)[32,40-41]、抗镰刀菌枯萎病[42]、抗条锈病[9]、抗叶锈病[43]、抗茎锈病[22]和抗白粉病基因[11,44-45]已经被鉴定并从中间偃麦草转移到小麦中应用。
白粉病和条锈病是两种世界最严重的小麦疾病,每年在全世界引起小麦大规模减产。已有报道中,中间偃麦草Js或St染色体上的白粉病和条锈病抗性基因已经转移运用到小麦中。B.Friebe等[43]报道了一个被定位在7Ai-2染色体上的条锈病抗性基因,Tang S.等[16]鉴定7Ai-2染色体属于Js染色体组。而来源于小麦-中间偃麦草后代品系X479的条锈病抗性基因则被定位在1St染色体上[12]。He R.L.等[44]研究发现白粉病抗性基因Pm43是从一条未知的Js染色体上转移到小麦的2DL上。通过C带和GISH鉴定,同样来源于普通小麦与中间偃麦草抗白粉病品系08-723,发现其携带的白粉病抗性基因来源于一条6AS与St的易位染色体[11]。在本研究中,品系12-1179包含6条St,4条Js和2条St-Js,对条锈病和白粉病具有免疫的特性,其白粉病和条锈病抗性基因应来源于St或Js。但其抗性基因是否与以往报道相同,还需要进一步研究。现在我们正在通过抗性筛选来选育具有抗白粉病与条锈病的外源附加系,代换系以及易位系小麦。
猜你喜欢
作物学报(2022年2期)2022-11-06
湖北植保(2022年4期)2022-08-23
今日农业(2021年9期)2021-11-26
今日农业(2021年7期)2021-07-28
快乐语文(2020年14期)2020-07-04
现代农村科技(2020年2期)2020-03-24
灾害学(2019年3期)2019-07-11
农民致富之友(2019年16期)2019-07-01
中国麻业科学(2018年6期)2018-04-09
西部(2016年6期)2016-05-14
