
12种真菌钾离子通道蛋白生物信息学分析
2018-09-13 11:09辛董董赵岩岩胡梁斌莫海珍李红波
食品工业科技 2018年16期
周 威,辛董董,赵岩岩,胡梁斌,莫海珍,李红波
(河南科技学院食品学院,河南新乡 453003)
真菌作为真核生物中一个庞大的类群,与人们的生活有着密切关系。香菇、木耳等大型真菌是人类的重要食物来源,米曲霉、酿酒酵母等是食品工业生产中重要的发酵菌种[1]。而黄曲霉、禾谷镰刀菌等病原性真菌却侵染粮食作物,危害食品安全、甚至人类健康[2-3]。抗真菌药物大量使用带来真菌耐药性日益严重,了解真菌耐药性机制、寻找新的抑制靶点有着重要的意义[4-5]。细胞内K+稳态是多数生物代谢机制和综合性能优化运行的先决条件。胞内K+动态平衡由细胞质和细胞膜上的许多K+通道和转运蛋白所调控[6]。在孢子萌发早期阶段,膜通透性增加,导致孢子内离子和水重新分配,孢子内80%的K+快速释放到孢子外,随后K+依赖能量被再次吸收[7]。而离子的移动往往涉及到离子通道的响应,这也是细胞识别外界信号的一个重要途径。钾离子通道蛋白在不同的真菌中分布广泛,目前已在酿酒酵母、米曲霉、烟曲霉等真菌中发现许多钾离子通道蛋白。钾离子通道是离子通道中种类最多。钾离子通道是目前存在最广泛且最复杂的
一大类钾离子通道,与多种生命活动密切相关,是药物设计的重要靶标之一[8]。对于钾离子通道结构和功能的研究主要通过实验的方法进行的,比如人工膜离子通道重建技术、细胞膜片钳和单通道记录技术、通道蛋白分离、基因重组技术、纯化等生化技术。在理论方面,对钾离子通道的研究主要有动力学模拟方法和序列分析方法。目前关于真菌的钾离子通道蛋白,研究较少。Gobert等[9]通过敲除和过表达拟南芥中钾离子通道TPK1,发现TPK1对保持胞内K+动态平衡起着重要作用,能够影响种子的萌发、生长和气孔运动。敲除拟南芥花粉管K+通道AKT6后,花粉管萌发率与野生型相比大幅度降低,并且AKT6对外界pH变化非常敏感[10]。随着对K+通道的研究,尚有一些问题有待深入探讨,比如亚基的功能、基因突变与疾病的不一致性等。由于K+通道自身的复杂性和多样性使得对一些K+结构和功能的认识上存在一定的局限性。多学科交叉研究和先进的技术方法的使用,对于探讨某些疾病或状态的机制、早期诊断及发现特异性治疗药物或措施等均具有十分重要的理论和实际意义[11]。
利用生物信息学对蛋白进行序列分析,从而推断并预测其结构和功能,已成为初步确定基因结构及功能的一种捷径[12]。本文筛选了酿酒酵母、黄曲霉、烟曲霉、禾谷镰刀菌、玉米黑粉菌、布拉克须霉等12种真菌的钾离子通道蛋白,采用生物信息学分析了这些蛋白的理化性质、结构特征和系统进化等,以期为真菌钾离子通道蛋白结构与生理功能研究奠定基础。
1 材料与方法
1.1 材料与仪器
12种真菌酿酒酵母(Saccharomycescerevisiae)、黄曲霉(Aspergillusflavus)、烟曲霉(A.fumigatus)、禾谷镰刀菌(Fusariumgraminearum)、玉米黑粉菌(A.flavus)、布拉克须霉(Phycomycesblakesleeanus)等。
NCBI数据库(http://www.ncbi.nlm.nih.gov/genbank/)。
1.2 实验方法
从NCBI数据库(http://www.ncbi.nlm.nih.gov/genbank/)中查找12种真菌钾离子通道蛋白的氨基酸序列(表1)。

表1 12种真菌钾离子通道氨基酸和核苷酸序列号Table 1 Amino acid and nucleotide sequence number of 12 fungal potassium channel proteins
利用ProtParam(http://www.expasy.org/tools/protparam.html)分析核酸及氨基酸序列的组成成分及理化性质。采用CLCCombinedWorkbench6观察图形化的多重序列比对结果。采用WebLogo产生序列logo图谱[13]。利用ClustalX1.83和MEGA6.0.5软件对12种真菌钾离子通道氨基酸和核苷酸序列构建系统进化树[14]。利用NPS分析蛋白质的二级结构[15]。利用SWISS-MODEL分析蛋白质的三级结构[16]。
利用HOMCOS进行同源四聚体结构模拟。
1.3 系统进化树的构建
应用多序列比对工具ClustalX1.83,以氨基酸全序列联配的结果为基础,用MEGA6.0.5软件中的邻接法(neighbor-joining,NJ)构建系统发育树,通过随机逐步比较的方法搜索最佳系统进化树。
2 结果与分析
2.1 真菌钾离子通道蛋白理化性质分析
利用ProtParam在线工具分析酿酒酵母、黄曲霉、烟曲霉、禾谷镰刀菌、玉米黑粉菌、布拉克须霉等12种真菌钾离子通道蛋白的核苷酸及其氨基酸序列。结果表明(见表2),氨基酸序列长度在659~928 aa之间,分子量在73970.53~101330.60 Da之间,理论等电点在5.45~9.54之间,亚细胞定位分析表明真菌钾离子通道蛋白主要定位在质膜、液泡上。亲水性/疏水性分析显示疏水区域明显大于亲水区域,说明真菌钾离子通道蛋白亲水性较强,为亲水性蛋白。蛋白磷酸化位点分析表明,不同真菌钾离子通道蛋白具有3~18个丝氨酸位点,其中多数蛋白具有相对保守的3个丝氨酸位点。

表2 12种真菌钾离子通道蛋白理化性质分析Table 2 Analysis of physical and chemical properties of 12 fungal potassium channel proteins
2.2 真菌钾离子通道蛋白同源性比较
利用NCBI Blast程序对选取的12种不同真菌钾离子通道蛋白序列进行同源性比对。结果表明,米曲霉菌钾离子通道蛋白(XP_001821104.1)与黄曲霉菌(XP_002376854.1)的同源性最高达99.85%,与黑曲霉菌(XP_001393797.2)、烟曲霉菌(XP_752795.1)的同源性为59.37%,与扩展青霉(XP_016598289.1)的同源性为51.69%,与油菜茎基溃疡病菌(XP_003842737.1)的同源性为38.13%,与灰葡萄孢菌(EMR91056.1)的同源性为36.43%,与禾谷镰刀菌(XP_011328543.1)的同源性为27%,与酿酒酵母(CAA64176.1)的同源性为19.54%,与玉米曲霉菌(XP_001821104.1)的同源性为15.36%,与布拉克须霉(XP_018296227.1)的同源性为15.04%,与长雌异水霉(KNE72472.1)的同源性为13.59%。
采用CLC Combined Workbench 6观察图形化的多重序列比对结果(图1),证实了上述同源性比对的结果。WebLogo基于多序列比对信息,把多序列的保守信息通过图形表示出来。每个logo由一系列氨基酸组成,在每一个序列位置上用总高度表示此位置上的序列保守性,用氨基酸字母的高度表示出现的频率[17]。分析表明不同真菌钾离子通道蛋白具有特殊的钾离子选择器保守结构TXGYGD(图2)。

图1 12种真菌钾离子通道蛋白氨基酸序列对比Fig.1 Alignment of potassium channel proteins amino acid sequences in 12 kinds of fungal
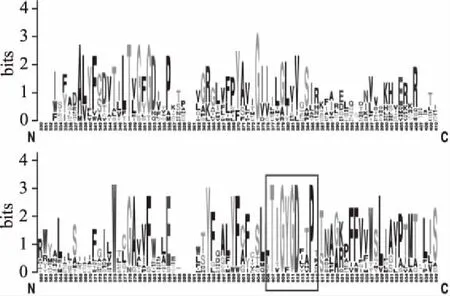
图2 12种真菌钾离子通道蛋白氨基酸序列Logo图谱Fig.2 The sequence logos of potassium channel proteins in 12 kinds of fungal by Web Logo
2.3 真菌钾离子通道蛋白系统进化树分析
真菌钾离子通道蛋白主要采用多序列比对工具ClustalX1.83和MEGA6.0.5软件。是以氨基酸全序列联配的结果为基础,采用软件中的MEGA6.0.5邻接法(neighbor-joining,NJ)构建系统发育树。对上述12种真菌钾离子通道蛋白氨基酸序列通过随机逐步比较的方法,搜索最佳系统进化树,对生成的系统树进行Bootstrap校正。构建系统进化树,采用默认参数。结果表明(图3),分属于担子菌门、子囊菌门、结合菌门以及壶菌门的各类真菌钾离子通道蛋白能够分别聚类在一起。其中,米曲霉(A.oryzae,XP_001821104.1)和黄曲霉(A.flavus,XP_002376854.1)在同一个进化分支上,同源性最高,其次为黑曲霉,这与同源性比较的结果相一致。

图3 12种真菌钾离子通道蛋白系统进化分析Fig.3 The phylogenetic analysis of potassium channel proteins in 12 kinds of fungal
2.4 真菌钾离子通道蛋白二级结构分析
蛋白质的二级结构是蛋白质分子的多肽链通常在一级结构的氨基酸折叠盘曲成比较稳定的,才能进一步完成具有蛋白质活性的三级或四级构象,以完成特定的生命活动。蛋白质的二级结构通常由α-螺旋、β-折叠、γ-转角、延伸链及无规则卷曲等组件组成。真菌钾离子通道蛋白二级结构主要是通过NPS程序对12种真菌钾离子通道蛋白序列进行二级结构分析表明,真菌钾离子通道蛋白均由α-螺旋、β-折叠、无规则卷曲和延伸链等结构元件组成,其中螺旋所占比例最高,在40.92%~52.24%之间,其次为无规则卷曲或延伸链(表3)。在不同真菌钾离子通道蛋白中,上述3种元件的比例和分布存在差异,例如,米曲霉的α-螺旋占 44.87%,延伸链占13.97%,无规则卷曲占41.16%,这暗示钾离子通道蛋白在不同真菌中可能具有独特的功能。
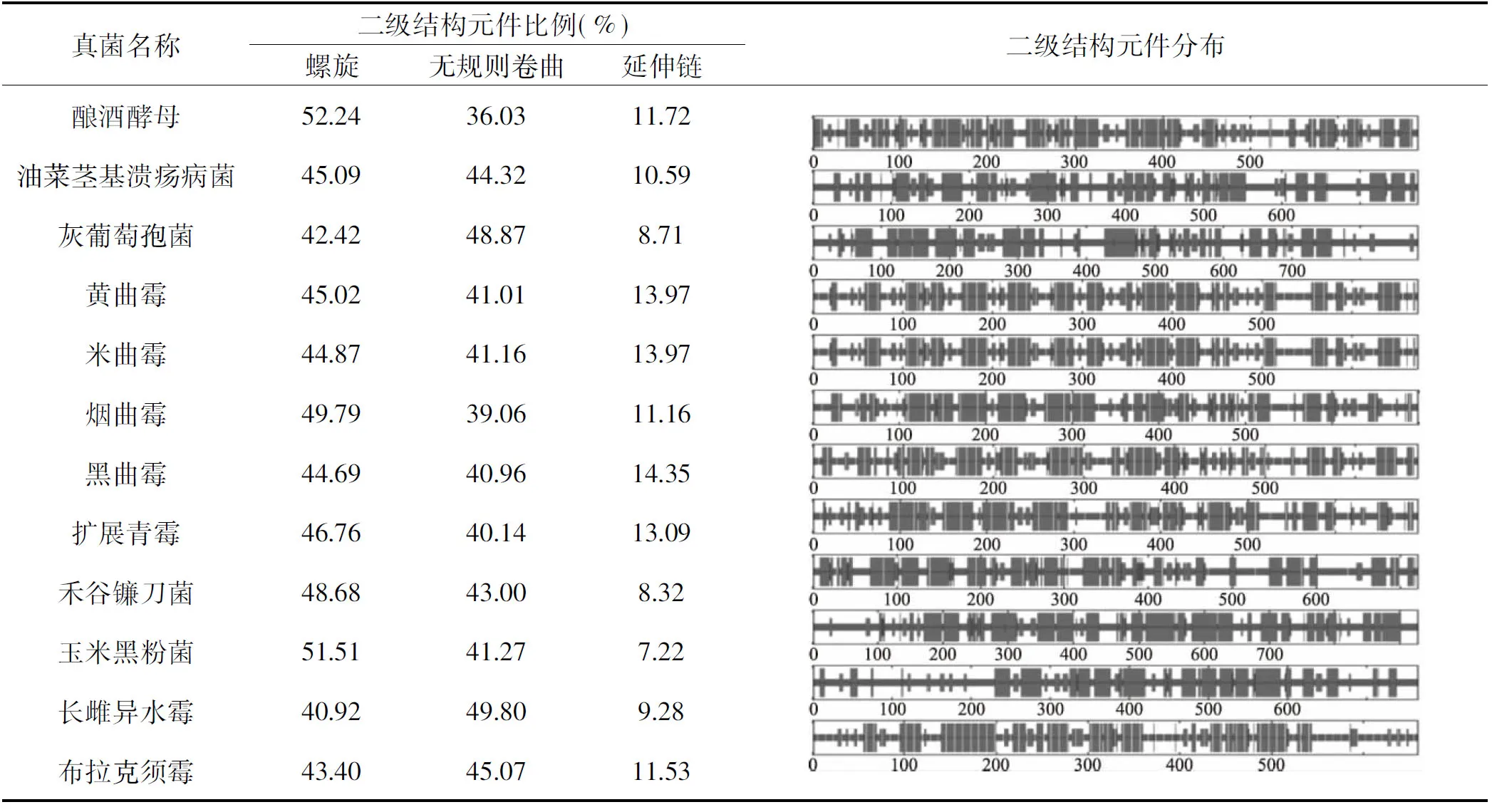
表3 12种真菌钾离子通道蛋白二级结构分析Table 3 The secondary structure analysis of 12 fungal potassium channel proteins
2.5 真菌钾离子通道蛋白三级结构分析
蛋白质的三级结构主要是指在二级结构的基础之上,经过再一次的充分折叠形成一个球形、包裹紧密、已经具有生物活性的完整三维立体结构,即天然构想[18]。利用Swiss-Model采用同源建模的方法预测不同真菌钾离子通道蛋白的三级结构。从图4可以看出,三级结构主要是由螺旋、无规则卷曲和延伸链等二级结构元件组成,其空间结构高度相似,具有高度保守的结构特征,表明它们可能具有相似的生物学功能。钾离子通道蛋白在活体膜中通常以四聚体结构存在,本文以黄曲霉菌钾离子通道蛋白(XP_002376854.1)为例,利用HOMCOS进行同源四聚体结构模拟,结果发现其符合钾离子通道结构特征,详见图5。不同真菌钾离子通道蛋白单体的拓扑结构和聚角度也存在差异,表明每种真菌钾离子通道蛋白可能具有自己独特的功能。
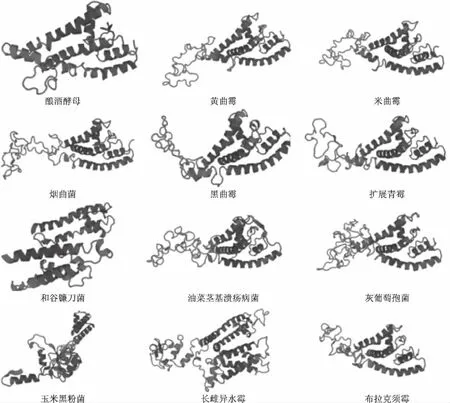
图4 12种真菌钾离子通道蛋白的三级结构分析Fig.4 The tertiary structure analysis of potassium channel proteins in 12 kinds of fungal
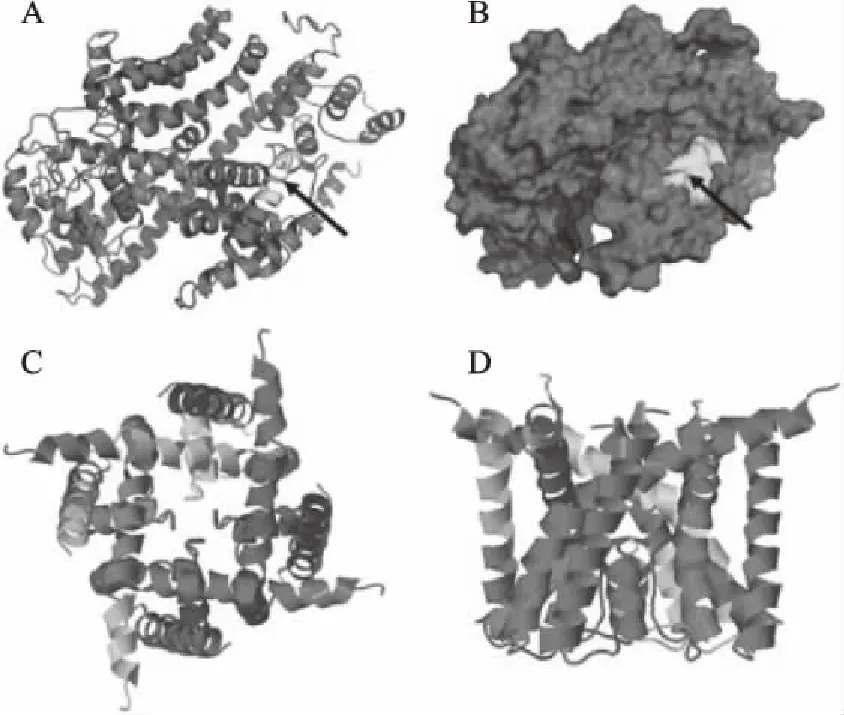
图5 黄曲霉菌钾离子通道蛋白的三级结构分析Fig.5 The tertiary structure analysis of potassium channel proteins in A.flavus
3 结论
本研究选择12种真菌钾离子通道蛋白进行生物信息学分析。一般认为,蛋白质的一级结构决定二级结构,二级结构决定三级结构。蛋白质的生物学功能在很大程度上取决于其空间结构,蛋白质结构构象多样性导致了不同的生物对环境适应性的不同。序列分析发现其都具有的保守结构域保守结构TXGYGD,属于钾离子通道钾离子选择器典型的特征片段。由二级结构分析表明,真菌钾离子通道蛋白均主由螺旋、无规则卷曲和延伸链等结构元件组成,但不同真菌中不同元件的比例和分布存在差异,这暗示钾离子通道蛋白在不同真菌中虽然具有相似的功能,但每种真菌的钾离子通道蛋白可能具有自身独特的功能。三维结构分析表明,黄曲霉菌钾离子通道蛋白在活体膜中是以四聚体结构存在的,这一结构对于蛋白质的结构稳定和功能行使的正确性有重要作用。蛋白质磷酸化是最重要的蛋白质翻译后修饰方式之一,可有效调控蛋白质活力和功能,并在细胞信号转导过程中起重要作用。研究表明,磷酸化是真菌钾离子通道蛋白活性调节的一种重要方式,很多真菌钾离子通道蛋白在生物体内都会发生磷酸化,而本研究中的磷酸化位点分析表明主要分布在丝氨酸上,丝氨酸位点一般3~18个。多个磷酸化位点的存在可能在植物胁迫和亚细胞定位中起着重要作用,可增强真菌钾离子通道蛋白调控途径的多样性。
然而关于真菌钾离子通道的相关文献较少,真菌孢子萌发过程中是否涉及到钾离子通道更是尚无研究报道。通过解析真菌钾离子通道的结构与功能,可以更加深入地了解其在真菌生长发育侵染等过程中的作用机制,为病原性真菌的防治提供新的思路。
猜你喜欢
世界科学技术-中医药现代化(2022年2期)2022-05-25
天津市教科院学报(2021年5期)2021-11-10
生物学通报(2021年9期)2021-07-01
世界科学技术-中医药现代化(2021年12期)2021-04-19
科学(2020年2期)2020-08-24
小学科学(学生版)(2018年8期)2018-08-21
现代园艺(2017年23期)2018-01-18
中国医学创新(2017年7期)2017-03-31
江苏农业科学(2016年8期)2017-02-15
微生物与感染(2015年5期)2015-12-08
