
SUPPLEMENTAL DESCRIPTION OF MYXOBOLUS HAICHENGENSIS CHEN, 1958(MYXOZOA: MYXOSPOREA) INFECTING THE GILLS OF ABBOTTINA RIVULARIS BASILEWAKY: MORPHOLOGICAL ANDMOLECULAR DATA
2018-09-13 07:37LIPengZHAOXinXIBingWenandXIEJun
水生生物学报 2018年5期
LI Peng, ZHAO Xin, XI Bing-Wen, and XIE Jun
(1. Wuxi Fisheries College, Nanjing Agricultural University, Wuxi 214081, China; 2. Key Laboratory of Freshwater Fisheries and Germplasm Resources Utilization, Ministry of Agriculture, Freshwater Fisheries Research Center, Chinese Academy of Fishery Sciences, Wuxi 214081, China)
Abstract: Myxobolus haichengensis Chen, 1958 forms numerous small plasmodia on the gill filaments of wild cyprinid Abbottina rivularis Basilewaky. The species described originally was lacking important characters, which made the accurate identification difficult. Here, we supplemented its characteristics with morphological and molecular data. Plasmodia of M. haichengensis are oval. Mature spores are ellipsoidal-shaped in frontal view and fusiform-shaped in lateral view, measuring (10.8±0.7) μm (10.1—11.5 μm) long, (8.1±0.5) μm (7.5—9.0 μm) wide, and (5.7±0.4) μm (5.2—9.0 μm) thick; two unequal polar capsule are pyriform with tapering anterior, large polar capsule averaging (4.7±0.5) μm (4.8—6.7 μm) long and (2.5±0.2) μm(3.2—4.3 μm) wide; small polar capsule averaging (4.4±0.2) μm (4.1—4.8 μm) long and (2.2±0.1) μm(2.0—2.5 μm) wide; polar filaments coil with four to five turns. The nuclear 18S rDNA sequence was obtained and deposited in GenBank (KY965936), and sequences alignment analyses revealed that M. haichengensis was most similar with the actinosporean Hexactinomyxon type 2 (AY162272, 97%) released from the freshwater tublificid oligochaete Limnodrilus hoffmeisteri.
Key words: Myxobolus haichengensis; Gill; Abbottina rivularis
Myxozoans are microscopic, multicellular, and obligate endoparasitic cnidarians that mostly infect fish, with some species parasitizing amphibians, reptiles, birds and mammals[1]. Until now, approximately 2200 myxosporean species classified in 60 genera have been reported over the world[2]. However, some of them were originally described a long time ago with incomplete data, which makes the current evaluation of species diversity difficult[3—5]. To settle the above conundrum, a comprehensive approach integrating morphological, histological (organ or tissue tropism), molecular (DNA sequence) data for characterizing myxosporean species has been widely accepted and implemented[6,7].
Abbottina rivularisBasilewaky (Cypriniformes:Cyprinidae), is a small benthopelagic freshwater cyprinid, and widely distributes in East Asia[8]. Up to now, 28 nominal myxosporean species were recorded fromA. rivularisand its congeneric species in China:2 ofChloromyxum, 3 ofZschokkella, 5 ofChloromyxu, and 18 ofMyxobolus[5]. However, most of them were lack of molecular data. During the recent parasitological investigation, numerous plasmodia were found on the gills ofA. rivularisin the Taihu Lake.Strict morphological comparisons showed that the spores were consistent withMyxobolus haichengen-sisChen, 1958. According to the literature,M. haichengensisChen, 1958 was first discovered on the gills ofA. rivularisandPelteobagrus fulvidraco, from the lower reach of the Liao River in Haicheng, Liaoning Province, China. The original morphological descriptions were somewhat ambiguous and incomplete,attributing to that the mature plasmodia and trophozoite were not observed. In the present study, we supplement the characteristics ofM. haichengensiswith morphological and 18S rDNA sequence data based on newly collected specimens.
1 Material and methods
1.1 Samples collection
In July 2016, twentyA. rivularis, body length of 73 —112 mm, were netted from the Lake Taihu,Wuxi, China (31°51′N, 120°23′E). All fishes were transported alive to the laboratory.
Gross examinations of all organs were conducted according to Lom and Dyková (1992)[9]. Numerous plasmodia containing myxospores were found on the gills ofA. rivularis. Five plasmodia were isolated separately from the infected gill filaments, squeezed on a clean slide to make wet-mount slide, and observed under light microscope Olympus CX-31 equipped with digital camera. The spores on the slide were collected and used for molecular analyses.Measurements were performed on 50 fresh mature spores. All measurements are given in micrometers(μm, mean±SD) unless otherwise indicated. Line drawings of the myxospores were made based on the digitized images.
1.2 DNA isolation and sequencing
Genomic DNA was extracted using a QIAamp®DNA Micro Kit (Qiagen, Germany) according to the manufacturer’s instructions. The 18S rDNA was amplified with primers ERI-B1 and ERI-B10[10]. The PCR products were purified using a Sangon Biotech®sanorep Column DNA Extraction kit. The purified product was cloned into the pMD18-T vector(TaKaRa Qingdao, China), and 5 positive clones were sequenced with an ABI 3100 Genetic Analyzed automated DNA sequencer (Applied Biosystems).
Sequences were assembled and inspected with SeqMan (Lasergen package; DNAStar Inc., Madison,WI). Assembled sequence was deposited in GenBank(accession number KY965936) and verified by BLAST search. 56 sequences with high similarity were downloaded from GenBank and aligned using Clustal X 1.8 program with defaulting setting[11]. The alignment was corrected manually using the MEGA7.0 software[12].
1.3 Phylogenetic analyses
Phylogenetic analyses were conducted with Bayesian inference (BI) and maximum likehood (ML)methods. The appropriate nucleotide substitution model (GTR+G) was estimated with the lowest BIC(Bayesian Information Criterion) scores with software MEGA 7.0. Nucleotide frequencies were estimated from the data (A=0.255, T=0.266, C=0.198,G=0.281), six rates of nucleotide substitution were[AC]=0.089, [AG]=0.250, [AT]=0.127, [CG]=0.070,[CT]=0.355, [GT]=0.107. The proportion of invariable site was 0.35, and the alpha value of gamma distribution parameter was 0.38. ML phylogenetic trees were inferred with MEGA 7.0, and branch support was computed with 1000 replicates of bootstrap analyses. Bayesian analyses were conducted in software MrBayes ver. 3.1.2 with parameter setting nruns=4,nst=6, rates=invagamma, ngen=5000000. Posterior probability values were used as support for the Bayesian topology.
2 Results
Numerous plasmodia were generally found in the gill filaments of 7 (35%) out of the 20 examinedAbbottina rivularis(Fig. 1). No other organs exa mined had plasmodia. Plasmodia were oval, measuring(0.6—1.1) mm in diameter (n=10) (Fig. 1).

图1 海城碘泡虫寄生于棒花鱼鳃Fig. 1 Myxobolus haichengensis Chen, 1958 were found in the gill filaments of Abbottina rivularis Basilewaky
2.1 Description of Myxobolus haichengensis
Fresh spores ellipsoidal-shaped in frontal view,and fusiform-shaped in lateral view, measuring 10.8±0.7 (10.1—11.5) μm long, 8.1±0.5 (7.5—9.0) μm wide (n=40), 5.7±0.4 (5.2—9.0) μm thick (n=10)(Fig. 2C, D; Fig. 3). Spore surface smooth, spore valves symmetrical. Two unequal polar capsules pyriform with an apophysis at the top end, and 4—5 turns of the polar filament visible inside (Fig. 2C). The large polar capsule 4.7±0.5 (4.8—6.7) μm long, and 2.5±0.2 (3.2—4.3) μm wide; the small polar capsule 4.4±0.2 (4.1—4.8) μm long, and 2.2±0.1 (2.0—2.5) μm wide (Fig. 2A—C).
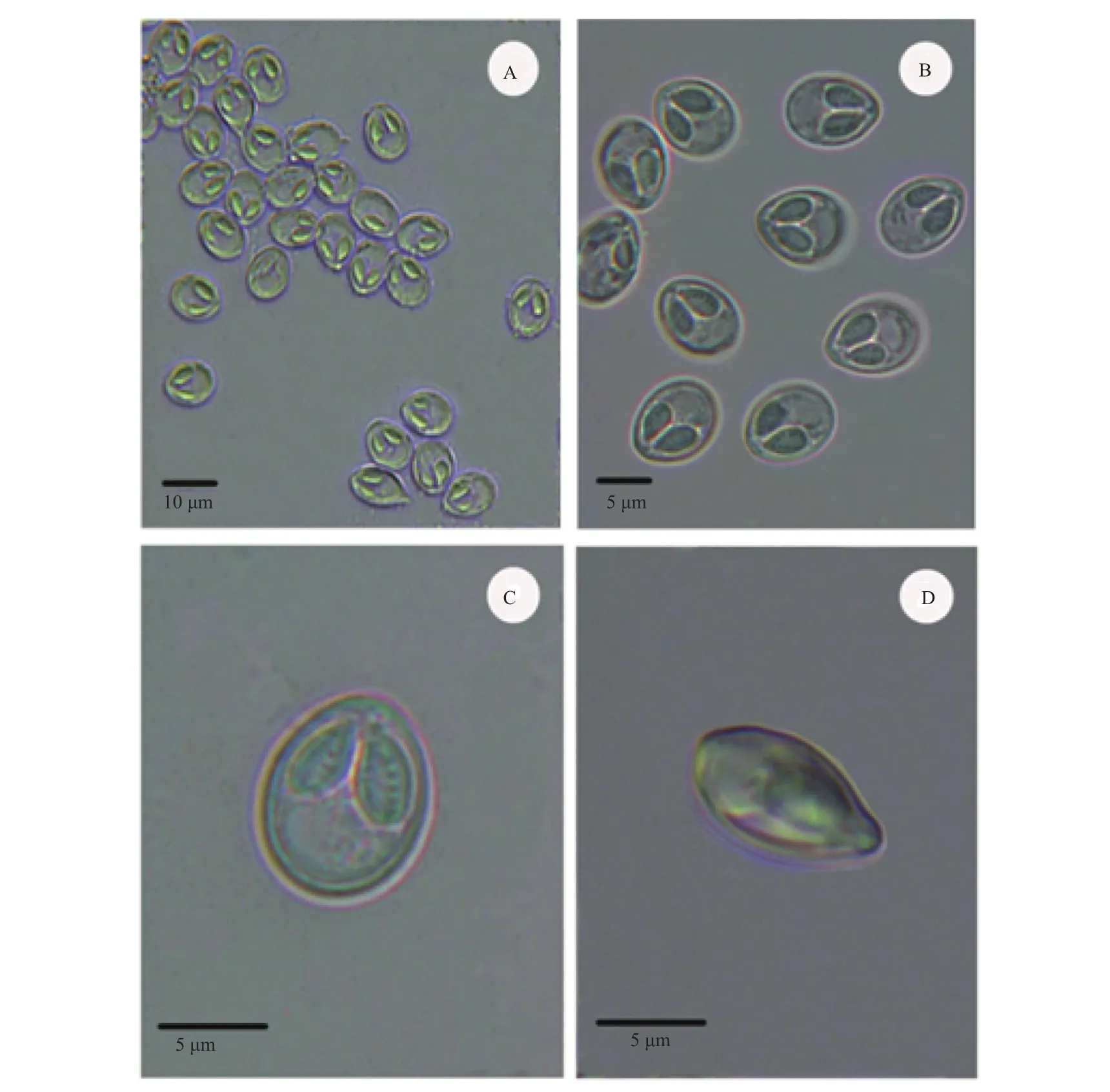
图2 海城碘泡虫显微镜下的孢子形态Fig. 2 Spores of Myxobolus haichengensis Chen, 1958 from Abbottina rivularis Basilewaky

图3 海城碘泡虫Fig. 3 Diagrammatic representation of Myxobolus haichengensis Chen, 1958 spores
2.2 Taxonomic summary
Type host:Abbottina rivularisBasilewaky (Cypriniformes: Cyprinidae)
Locality: The Taihu Lake (31°51′N, 120°23′E),Jiangsu Province; The Liao River, Haicheng, (type locality).
Site of infection: Gill filaments
Date of sampling: July 2016
Host size: 73—112 mm
Prevalence: 7 of 20 (35%)Abbottina rivularis.
Type material: Syntype specimens of spores in glycerin gelatin had been deposited in the Key Laboratory of Freshwater Fisheries and Germplasm Resources Utilization, Ministry of Agriculture, Freshwater Fisheries Research Center, Chinese Academy of Fishery Sciences (accession No. MTR20160715).
2.3 Molecular analyses
The partial SSU rDNA sequence was deposited in GenBank (2044 bp, accession No. KY965936).BLAST analyses showed that the most closely related sequences were that of actinosporeans Hexactinomyxon type 2 (AY162272, 97%), Hexactinomyxon type 1 (AY162271, 95%), and Hexactinomyxon type SH-2006 (DQ473517, 93%). However, the homologous sequences of myxosporeans obtained from fish available in GenBank present low than 93% sequence similarity.
Phylogenetic analyses based on 18S rDNA sequences of myxozoans revealed thatM. haichengensiswas firmly clustered in the clade consisted of actinosporeans Hexactinomyxon type 2, and Hexactinomyxon type 1, collected from freshwater tublificid oligochaetes,Limnodrilus hoffmeisteriandL.udekemianus, and Hexactinomyxon type SH-2006 from oligochaetePsammoryctides albicola.Myxobolus pfeifferiisolated from the gill arches of Iberian barbellLuciobarbus bocagei,Myxobolus caudatusfrom the fin and gill of common barbellBarbus barbus, andMyxobolus squamaefrom scales of common barbellBarbus barbus(Fig. 4).
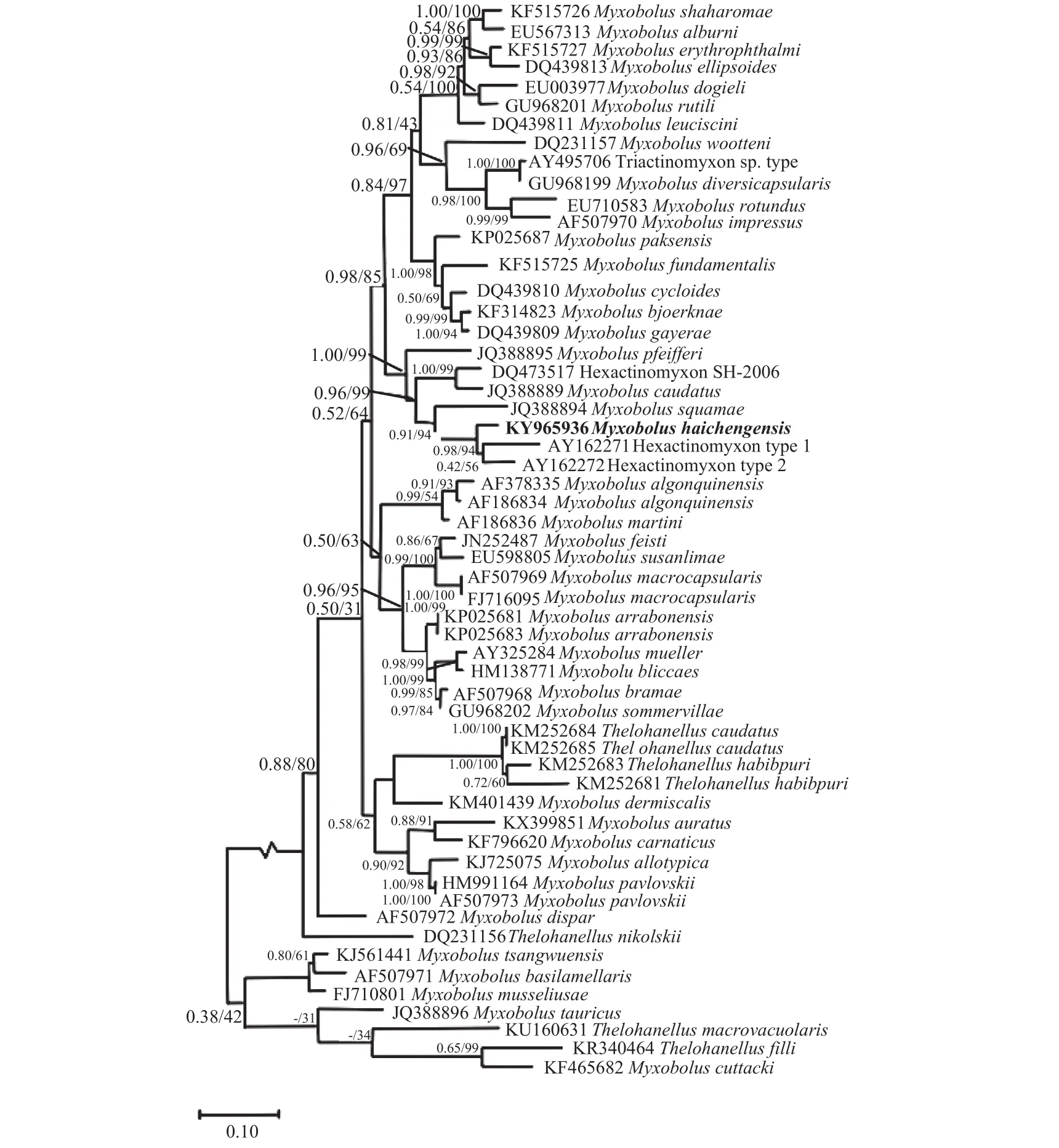
图4 基于18S rDNA基因序列构建的黏孢子虫贝叶斯法(BI)系统发育树Fig. 4 Phylogenetic tree generated by Bayesian analysis (BI) of the aligned SSU rRNA gene sequences of Myxobolus haichengensis Chen,1958 and related myxosporeans
3 Discussion
MyxobolusBütschli, 1882 is a species-rich genus among myxozoans, comprising 850 species throughout the world[13]. Among them, 18 species infecting different organs ofA. rivularisand congeneric species were previously recorded in the monograph of Chen and Ma (1998)[5]. In the original description,Myxobolus haichengensiswas described as a new species based to some fresh spore scattered on the gills. However, many important features were not pointed out, such as the size and shape of plasmodia,and the prevalence in host population. The present specimen ofM. haichengensishas similar spore morphological characters as the original description, expect the ratio of large to small polar capsule length(1.1vs. 1.0). Further comparison with the related species in literature revealed thatM. haichengensisis a valid species, and resemblesMyxobolus abbottiaeMa Dong & Wang 1982,Myxobolus kiatingensisMa 1979 andMyxobolus opsariichthysiLi & Nie 1973(Tab. 1). However,M. abbottiaecan be distinguished from the species reported here by having a higherratio of spore length to width (0.4vs. 0.6), and the former had a bigger spore size.M. haichengensiscan be readily distinguished fromM. opsariichthysiwith two unequal polar capsules, and different ratio of spore length to width (1.33vs. 1.28). It differed fromM. kiatingensisby having a different shape of anterior end of spores (roundvs. tapered), and ratio of spore thick to width (0.7vs. 0.9).

表1 海城碘泡虫与相似种类形态学比较Tab. 1 Morphological comparison of spore of Myxobolus haichengensis Chen, 1958 with related species
Myxozoan possesses a complex life cycle with two alternative stage, actinosporean developing in an invertebrate host and myxosporean developing in a vertebrate host. Recently, DNA sequences analyses have been widely used to identify the myxosporeanactinosporean counterparts (e.g. Ruidisch,et al. 1991;Rangel,et al. 2015; Xi,et al. 2015)[14—16]. In this study, the sequences alignment of 18S rDNA, showedM. haichengensiswas most closely related to actinosporeans Hexactinomyxon type 2 (97%), Hexactinomyxon type 1 (95%), and Hexactinomyxon type SH-2006(93%). The high sequence similarity indicated that these myxozoans were closely related species. Therefore, the life cycle ofM. haichengensismight involve a hexactinomyxon-type actinosporean.Phylogenetic analyses revealed thatM. haichengensiswas firmly clustered withMyxobolus pfeifferi,Myxobolus caudatusandMyxobolus squamaethat were collected from common barbell and Iberian barbell from Europe (Fig. 4). They had a common ancestor and possessed similar spore morphology, although occurred in different biogeographic regions.Furthermore, this lineage represented diverse sites of sporulation (e.g.M. pfeifferidwelling in the muscles and gill arch,M. caudatusin the fins and scales,M.squamaein the scales). Why thoseMyxobolusspecies distributing far apart and infecting not related fish host, showed a close phylogenetic relationship? Further studies were needed to reveal the evolution process.
猜你喜欢
中国科学探险(2022年7期)2022-12-30
Chinese Physics B(2022年9期)2022-09-24
导航定位学报(2022年4期)2022-08-15
音乐天地(音乐创作版)(2021年8期)2021-12-01
诗林(2020年1期)2020-11-18
发明与创新·中学生(2019年7期)2019-07-21
中国继续医学教育(2015年2期)2016-01-06
中学语文(2015年2期)2015-08-15
创新作文(5-6年级)(2014年12期)2014-12-19
振动、测试与诊断(2014年6期)2014-03-01
