
油菜MADS-box家族基因AGL13表达分析及RNA干扰载体的构建
2018-10-15 07:54熊书孙厚良杨丽珊马强李国利周大祥
生物学杂志 2018年5期
熊书, 孙厚良, 杨丽珊, 马强, 李国利, 周大祥
(1. 重庆三峡医药高等专科学校 基础医学部, 万州 404120; 2. 重庆三峡学院 生命科学与工程学院, 万州 404100)
作为一类重要的转录因子,MADS-box家族基因在真核生物中分布广泛[1]。该家族基因AGLs系列不仅参与植物花器官的发育[2-3],而且在果实的发育过程中也扮演着重要的作用[4-5]。
研究已发现,多种转录因子在影响种子发育的同时,对油脂的合成和积累具有重要的调控作用,如Fusca3(FUS3)、 ABI3 (ABSCISIC ACID INSENSITIVE3)、Leafy Cotyledon1,2(LEC1,LEC2)、WRINKLEDl(WRI1)和Dof (DNA binding with One Finger)等。这些基因的突变、过量表达或异位表达都会对植物种子发育和油脂等物质的积累产生重要的影响。FUS3作为一种转录因子调控种子发育及参与油脂积累,对甘蓝型油菜FUS3突变体的研究发现,抑制种子油脂合成导致蛋白质含量增加,参与脂肪酸修饰、蔗糖代谢和糖酵解的基因在突变体植株的发育种子中被下调表达[6]。在拟南芥中表达蓖麻的LEC2基因可使脂肪酸链延长酶1表达上调,同时诱导甘油三酯的积累[7]。而衰老诱导拟南芥LEC2使脂肪酸和甘油三酯生物合成的相关基因上调表达,进而显著提高叶部C20:1和C22:1脂肪酸的合成[8]。在种子发育早期,WRI1调控脂肪酸的从头合成及糖酵解过程,完成了种子中糖代谢向脂肪酸代谢的转化,在种子发育中期通过延长油脂生物合成可以提高拟南芥种子的油脂含量[9-10]。在莱茵衣藻中,超表达Dof型转录因子可增加转基因藻中油脂的积累[11]。
通过油菜胚发育和油脂合成表达数据库,我们发现AGL13基因在胚中表达较高,初步认为该基因可能与油脂合成有关。本研究拟克隆甘蓝型油菜(BrassicanapusL.)的AGL13基因,同时构建AGL13的干扰载体,为后续实验进一步确定AGL13通过控制油脂合成相关基因的表达,从而调节油脂的合成,线索也为通过基因工程手段提高油菜油脂含量,改良油菜品质奠定基础。
1 材料与方法
1.1 材料
以Brassicanapuscv. Westar甘蓝型油菜作为研究材料,实验所用的RNAi干扰质粒和农杆菌菌株GV3101由重庆大学基因工程中心提供。
1.2 方法
1.2.1 提取RNA与荧光定量PCR
以BnAGL13基因设计荧光定量PCR引物AGL13-F:5′-GGAAGAGGGAGAGTGGAGAT-3′;AGL13-R:5′-TCAATTGTTCTTTCAACTCC-3′,引物由华大基因合成。利用OMEGA植物RNA提取试剂盒提取油菜各部位的RNA,利用Thermo Scientific First Strand cDNA Synthesis Kit试剂盒反转录获得cDNA,-80℃保存备用。荧光定量PCR利用TaKaRa公司SYBR Green Ⅰ试剂盒,在CFX96荧光定量PCR仪上进行,分析BnAGL13基因在油菜不同组织的表达情况。反应程序:95℃预变性20 s;95℃变性3 s,60℃退火30 s,60℃延伸15 s,39个循环。每个样品5次重复,数据采用2—△△Ct法计算。
1.2.2 生物信息学分析
序列信息在NCBI数据库下载,蛋白结构域分析在http://pfam.wustl.edu/上进行。利用ClustalX2.1比对蛋白质序列,用MEGA 5构建进化树, 构树方法为Construct/Test Neighbor-Joining Tree,利用SWISS MODEL(http://swiss model.expase.org)预测蛋白质的三维结构,采用First approach mode建模,参考拟南芥的晶体模型。
1.2.3 干扰载体构建
以油菜盛花后5 d的种子cDNA为模板,用BnAGL13的正向前后引物(For-BnAGL13-F: 5′-CCGCT
CGAGCGCAAGACACAAGTTATGAT-3′;For-BnAGL13-R: 5′-GGGGTACCGTAAAAAGGGTTCGGTGTTG-3′)扩增出正向基因片段For-BnAGL13,将此片段以及RNAi载体用Xhol和Kpn1酶切,纯化回收后连接到RNAi载体上,转化到大肠杆菌,酶切验证。反向前后引物(Rev-BnAGL13-F: 5′-GCTCTAGAGTAAAAAGGGTTCGGTGTTGC-3′; Rev-BnAGL13-R: 5′-CCCAAGCTTCGCAAGACACAAGTTATGAT-3′)操作同上,用XbaI和HindIII酶切载体。
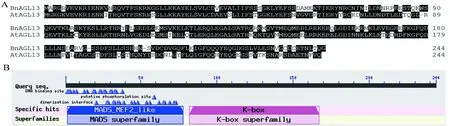
At:拟南芥;Bn:甘蓝型油菜
图1 AGL13的氨基酸序列比对和同源性分析
Fig 1 Amino acid sequence alignment and homology analysis ofAGL13
1.3 数据分析
利用统计软件SPSS19.0对试验数据进行t检验,P<0.05说明差异显著。
2 结果与分析
2.1 蛋白结构分析
将甘蓝型油菜与拟南芥的AGL13蛋白结构进行比对分析,我们发现油菜的AGL13氨基酸序列与拟南芥同源性很高,二者都具有完整且典型的MADS-box保守结构域(图1),对该蛋白的结构功能位点进行预测,发现该蛋白具有DNA结合位点,位于C端区域。许多研究发现,MADS-box家族基因在植物花和果实的发育过程中起着重要的作用,我们推测AGL13蛋白编码的基因属于MADS-box家族转录调控因子。
2.2 蛋白质的三维结构预测
SWISS MODEL在线分析蛋白质三级结构,我们发现该蛋白为同源四聚体结构,具有两个α螺旋,螺旋之间为无规则卷曲,其整体形态呈现典型的DNA核酸结合结构(图2)。表明该基因表达的蛋白质为转录因子,具备典型的MADS-box保守结构域,是MADS-box家族基因。

图 2 SWISS MODEL三维蛋白质结构预测图
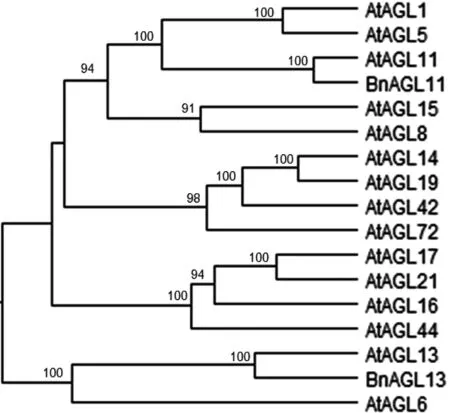
图3 BnAGL13的系统发育树
2.3 进化树分析
从NCBI数据库下载蛋白序列,利用MEGA程序构建系统发育树,分析BnAGL13蛋白和AtAGL蛋白的遗传进化关系。从图3可知,BnAGL13蛋白与AtAGL13和AtAGL6蛋白组成一个小的亚家族,其中,BnAGL13蛋白与AtAGL13蛋白亲源关系最近,说明BnAGL13可能与AtAGL13在功能上可能有一定的相似性。
2.4 定量PCR表达分析
我们分析了油菜各组织AGL13基因的组织表达模式,结果发现AGL13在各组织中均表达,其中在茎部表达水平最低,果夹15DAP时期表达水平最高,盛花期到种子45DAP表达水平都很高(图4),在果夹5DAP和果夹15DAP之间是油菜种子胚迅速膨大、油脂快速积累的时期,定量PCR的结果表明AGL13可能与油菜种子发育及油脂的合成密切相关。
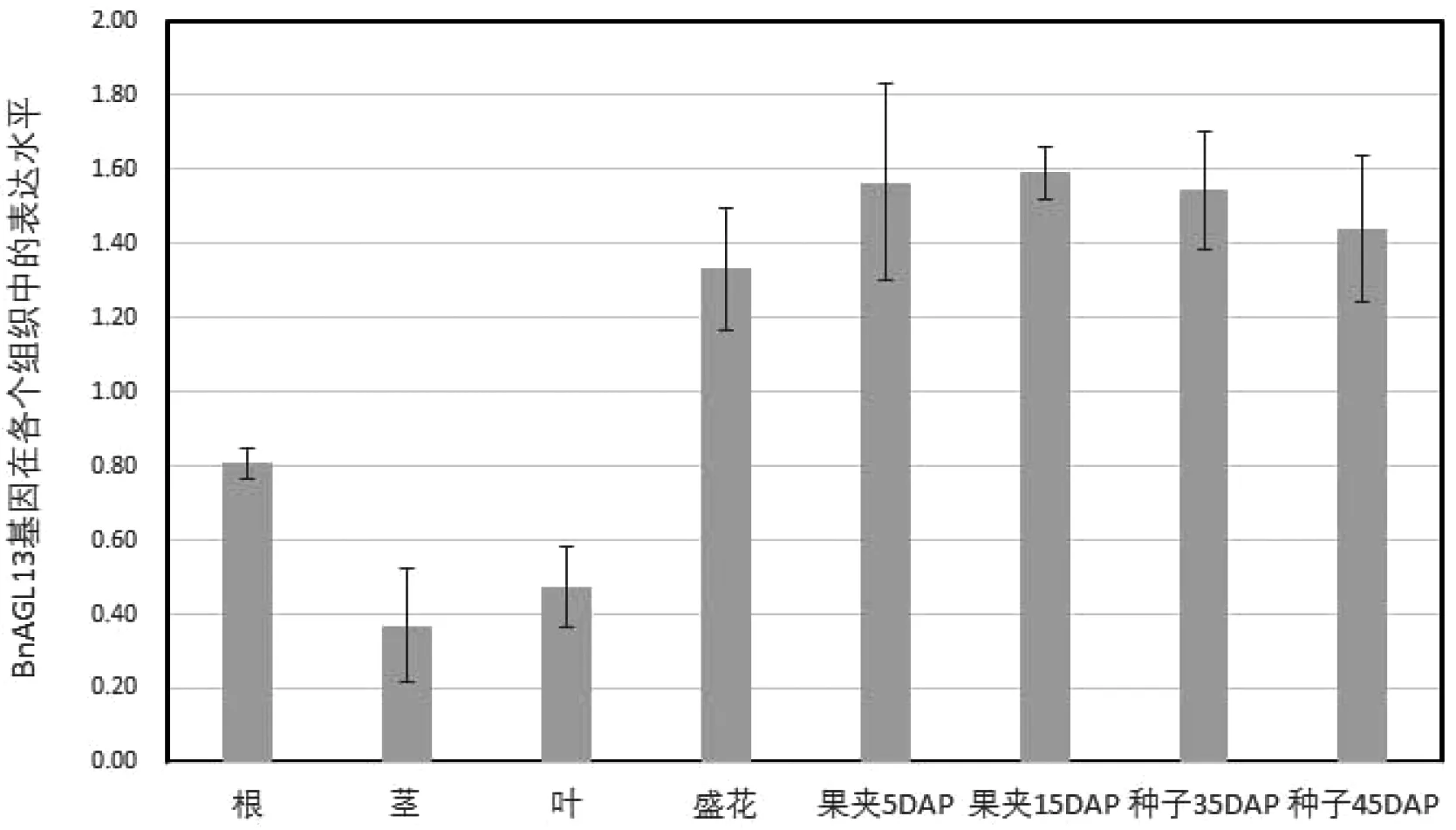
DAP:盛花后的种子
图4甘蓝型油菜AGL13基因的组织表达模式分析
Fig 4 Analysis of tissue expression patterns ofBnAGL13 gene
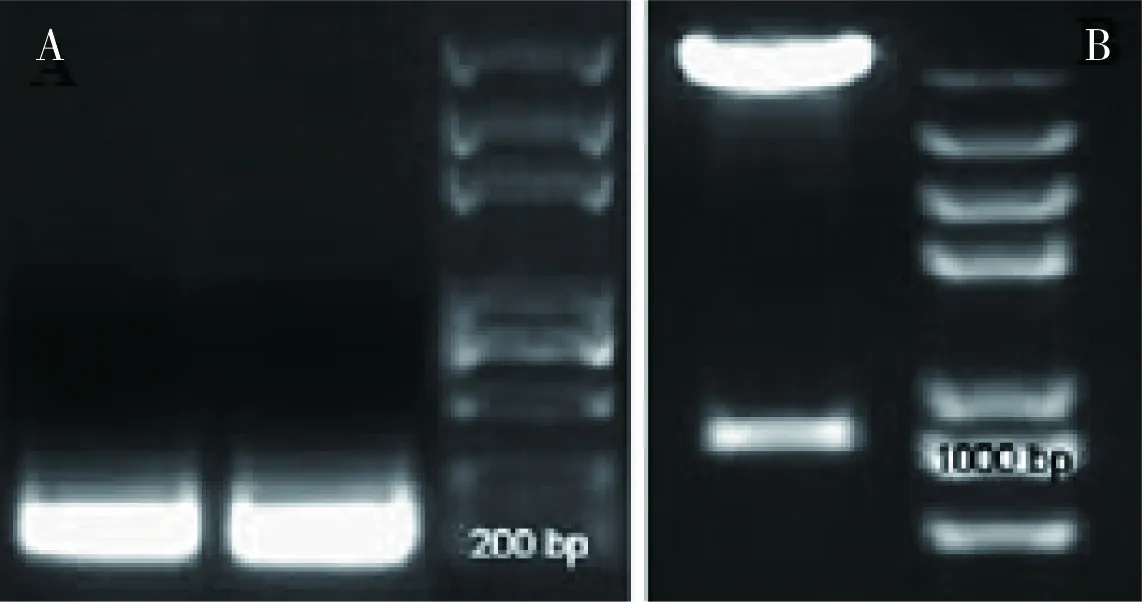
图5 RNAi干扰载体的构建
2.5 干扰载体构建及转化油菜
为了进一步验证AGL13在油菜种子发育及其油脂合成中可能具有重要的作用,我们采用RNA干扰技术下调AGL13基因在油菜中的表达,观察转基因油菜的表型。在构建RNA干扰载体时,我们首先以AGL13基因的cDNA作为模板,200 bp的干扰载体正向和反向片段分别被扩增出来(图5-A),然后分别插入干扰载体中,获得重组质粒,进一步酶切验证插入片段大小,发现插入片段为1000 bp(图5-B),与预期一致,证明RNAi载体已经构建成功。下一步我们将构建好的载体导入农杆菌GV310菌株中,采用农杆菌介导的叶盘法,用油菜下胚轴和子叶为受体,进行遗传转化,目前正在获得阳性抗性苗。随后将进一步获得转基因油菜植株,观察转基因油菜胚的发育和种子油脂的积累,确定AGL13在油菜胚的发育及油脂的合成中发挥着重要的作用。
3 讨论
甘蓝型油菜油广泛应用于人类消费和工业生产,对更新、污染更小和廉价燃料资源的需求增加了对生物燃料生产的探索。有数据显示,我国菜籽油占食用植物油使用量的三分之一,而菜籽饼粕占植物饼粕使用量的四分之一,增加油菜产油率和油脂品质改善始终是油菜育种的重要内容。对单位面积产油量而言,油菜油脂含量每提高1%,相当于增产菜籽2.5%。目前,我国主要的油菜品种含有40%左右的平均油脂含量,而国外一般油菜品种在42%~45%之间,部分甘蓝型油菜品种油脂含量高达47%[12]。因此,我国油菜种植业经济效益低下,这必然会打击农民种植油菜的积极性,制约了进一步扩大油菜种植面积和油菜业的发展。为此,进一步研究油菜油脂合成的分子调控机理,可以让我们了解控制油脂合成的关键基因,为通过基因工程手段提高油菜油脂含量,改良油菜品质,提高油菜产业生产效益奠定基础。
多种转录因子在影响种子发育的同时,对油脂的合成和积累具有重要的调控作用。研究发现,拟南芥LEC2突变体改变了种子的化学组成,降低了油脂和蛋白质含量,而淀粉和蔗糖含量反而增加[13]。在油菜中WRI1转录因子促进植物开花,增强种子和叶组织的油脂积累,对植物生长没有明显的抑制作用。该转录因子是通过降低贮藏的碳水化合物,增加可溶性糖来促进脂质代谢的碳通量,调节油脂积累和膜脂代谢之间的平衡[14]。而拟南芥种子发育中期,超表达WRI1将延长油脂生物合成,提高油脂含量,且种子中油脂含量与蛋白质含量呈负相关[15]。TTG1作为WRKY家族的一员,在调节拟南芥种子储备营养成分中起着重要的作用,FUS3可直接结合到TTG1基因组区域中,抑制种子的TTG1表达,从而调控脂肪酸成份及含量[16]。
定量PCR实验证明了AGL13基因在种子发育及油脂积累过程中表达强烈,表明该基因可能在这些发育过程中发挥着重要作用。后续实验将通过获得抑制AGL13基因表达的转基因油菜,并对转基因油菜进行胚分析和油脂含量测定,观察转基因植株的种子及胚发育是否被强烈抑制,胚中的脂肪酸的含量是否显著降低。通过上述研究,明确了AGL13基因在控制油菜种子中脂肪酸合成中的作用。
猜你喜欢
国际放射医学核医学杂志(2020年4期)2020-07-27
疯狂英语·新读写(2020年3期)2020-06-06
西藏农业科技(2019年3期)2019-11-04
西藏农业科技(2019年1期)2019-07-25
中国粮油学报(2019年4期)2019-07-12
食品与机械(2018年11期)2019-01-03
西藏农业科技(2018年4期)2018-04-25
中国粮油学报(2018年12期)2018-01-26
上海农业学报(2017年3期)2017-04-10
天然产物研究与开发(2014年6期)2014-04-27
