
冬季覆盖作物秸秆还田对双季稻田根际土壤微生物群落功能多样性的影响
2018-11-02 06:36唐海明肖小平汤文光郭立君孙玉桃程凯凯潘孝晨
生态学报 2018年18期
唐海明,肖小平,李 超,汤文光,郭立君,汪 柯,孙玉桃,程凯凯,孙 耿,潘孝晨
湖南省土壤肥料研究所, 长沙 410125
微生物是土壤生物中最重要的组成部分,它是土壤环境质量重要的指标,在土壤理化特性、生态环境、作物生长等方面具有十分重要的意义[1]。土壤微生物群落功能多样性能反映土壤中微生物的生态特征,它是土壤微生物群落状态与功能重要的指标。目前,测定土壤微生物群落功能多样性的方法较多,其中,Biolog微平板分析法是测定土壤微生物功能和结构多样性较为普遍的方法[2];该种方法是通过测试微生物对单一能源碳利用程度,来反映微生物群体水平的生理轮廓[3]。常用的方法是用Biolog-GN板进行碳源的测试,该方法基本能测定所有能源碳的种类[4];且操作简单,获得数据量丰富,能反映微生物种群的总体活性,近十几年在微生物生态及环境微生物检测方面应用均十分广泛[5]。
土壤微生物群落功能多样性易受稻田生态环境变化的影响,近年来,大量的学者主要开展了不同种植制度、作物类型和品种、土地利用方式、土壤类型、土壤理化特性、施肥模式、秸秆还田、土壤耕作、水分管理等条件下对土壤微生物群落结构和功能多样性影响的研究,Buenemann等[6]研究结果表明,不同的种植制度和作物类型对土壤微生物群落结构和功能多样性均具有显著的影响。张杰等[7]开展不同土地利用方式下土壤微生物群落功能多样性,结果表明退田还湖有利于恢复湿地土壤微生物群落结构。时鹏等[8]研究认为,玉米连作配施秸秆和撂荒均有利于提高土壤微生物代谢活性和土壤微生物群落功能多样性。Garland等[2]研究结果表明,不同土壤类型的土壤微生物群落结构存在明显的差异。Hu等[9]研究表明,长期平衡施肥使土壤微生物量碳、氮和微生物功能活性增强。侯晓杰等[10]认为,合理施用肥料有利于增强土壤碳源平均颜色变化率(AWCD),显著增加微生物功能多样性。徐一兰等[11]、罗希茜等[12]和徐万里等[13]认为,有机肥与无机肥配施均有利于提高土壤微生物多样性。周文新等[14]研究结果表明,67%和100%稻草还田措施对土壤微生物功能多样性具有明显的增加作用,其中以67%稻草还田处理的效果为最佳。然而有研究结果表明,长期施用无机氮肥降低了土壤微生物活性[15]。
湖南是我国主要的双季稻区,保持该区域稻田土壤肥力的稳定和持续是水稻高产、稳产的重要基础,对于保障国家粮食生产安全具有重要的战略意义。农田冬季覆盖作物是农业可持续发展的重要组成部分,据不完全统计,我国南方稻区约有冬闲田2.0×107hm2,利用冬闲田发展冬季农业,其中油菜(BrassicanapusL.)5.89×106hm2、紫云英(AstragalussinicusL.)1.86×106hm2、马铃薯(SolanumtuberosumL.)1.22×106hm2和黑麦草(LoliummultiflorumL.)7.67×105hm2 [16]。在该区域冬季具有优越的温、光、水、土等自然资源,若能充分利用丰富的自然资源和冬闲田开展冬季农业生产,可增加冬季稻田绿色作物覆盖度,有利于减少农田冬季裸露和土壤流失、增加碳氮蓄积、改善土壤理化性状,从而增加后茬水稻产量和保持农田的可持续发展[17-18]。目前,在该区域对不同冬季覆盖作物秸秆还田后对双季稻田土壤养分、土壤部分理化特性、温室气体排放、后茬水稻生物学和理化特性及水稻产量等方面影响进行了相应的研究[19-23],但关于冬季覆盖作物秸秆还田后对早、晚稻成熟期稻田根际土壤微生物群落功能多样性的影响还有待进一步开展。因此,本研究选择南方双季稻区4种具有代表性的冬季作物,以冬闲-双季稻种植模式为对照,利用Biolog-GN技术,初步探讨了黑麦草、紫云英、马铃薯和油菜-双季稻4种种植模式条件下双季稻田水稻根际土壤微生物功能的响应,从而为科学评价不同冬季覆盖作物-双季稻高效种植模式的可持续发展提供理论依据。
1 材料与方法
1.1 试验地概况
试验在湖南省华容县三封寺镇群力村试验基地进行。试验土壤为紫潮泥。试验开始于2014年10月,试验前耕层土壤基础养分性状为:有机质30.96 g/kg,全氮2.01 g/kg,全磷0.48 g/kg,全钾17.7 g/kg,碱解氮166.6 mg/kg,有效磷10.04 mg/kg,速效钾72.0 mg/kg,pH值6.20。试验地属湿润性大陆季风气候,年均气温17.0—18.0 ℃,年均降水量1200 mm,≥10 ℃活动积温5000—5600℃,无霜期260 d。
1.2 试验设计及田间管理
试验设5个处理:冬闲-双季稻(CK)、黑麦草-双季稻(Ry)、紫云英-双季稻(Mv)、油菜-双季稻(Ra)和马铃薯-双季稻(Po),每处理3次重复,随机区组排列,小区面积50.0 m2,规格为20.0 m(长)×2.5 m(宽)。不同小区间用塑料薄膜包埂隔离,保证单独肥水管理;冬季作物和水稻生育期具体的肥料用量及田间管理措施见表1。早稻和晚稻生育期,稻田水分管理采用前期浅薄灌溉、中期烤田和后期干湿交替的管理模式,其他管理措施同常规大田生产。每年的施肥方法和田间管理措施如灌溉、使用机械、喷洒农药等均一致。
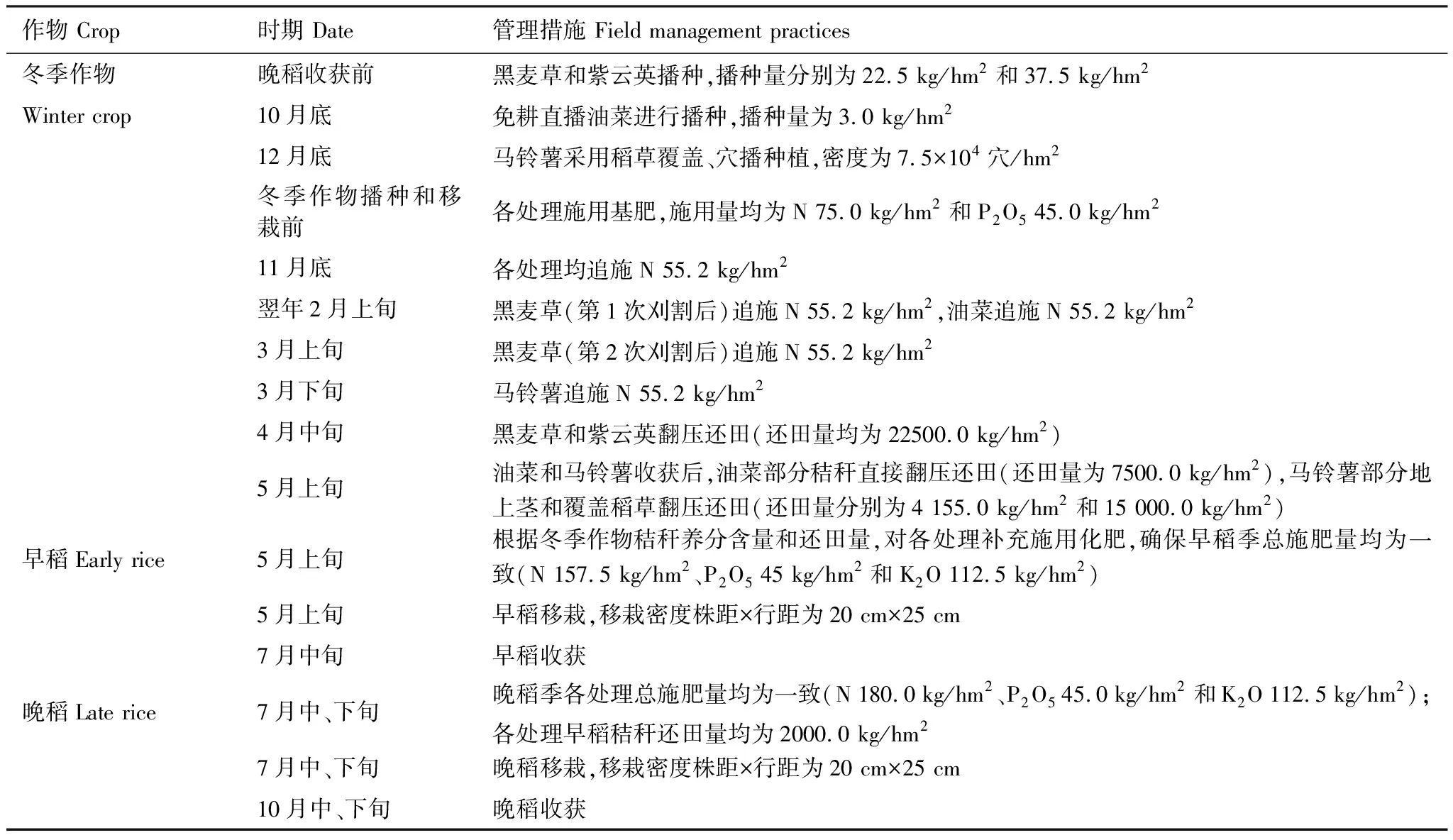
表1 双季稻田冬季作物和水稻生长期田间管理措施
1.3 样品采集与测定方法
2017年,分别于早稻成熟期(7月15日)和晚稻成熟期(10月17日)进行土壤样品的采集,每个小区采取多点取样法采集水稻根际土壤组成一个样品,重复3次。根际土壤取样方法采用“抖根法”,即先将植株根系从晒田后的稻田土壤中挖出,抖掉与根系松散结合的土体,然后将与根系紧密结合在0—4 mm范围的土壤用刷子刷下来作为根际土壤[24]。剔除土壤中石砾及植物残茬等杂物后,迅速装入塑料袋并放置于冰盒中,运至实验室,4℃冰箱保存,Biolog-GN分析在取样后48 h内进行。同时,在晚稻成熟期收获时采集各处理0—20 cm土壤新鲜样品,风干过筛,用于测定土壤pH值(土∶水=1∶2.5)、有机质(重铬酸钾容量法)、全氮(重铬酸钾-硫酸消化法)、碱解氮(碱解扩散法)、有效磷(NaHCO3浸提-钼锑抗比色法)以及速效钾含量(NH4Ac浸提-火焰光度法)[25]。
试验使用的Biolog-GN测试板共96孔。测试板的第1个孔为不含任何C源的对照,其余每孔中含有1种C源和氧化还原染料四氮唑蓝。微生物利用碳源进行呼吸使氧化还原电势发生变化,并将四唑类(TV)从无色还原成紫色[26]。通过测定各板孔的吸光值及其变化来反映微生物群落代谢功能的多样性。具体实验步骤是,取经预培养(26℃下培养4—6 h)的新鲜土样10 g,加95 mL无菌的0.145 mol/L NaCl溶液在摇床上振荡15 min(5 g土壤+95 mL无菌水),然后用无菌水分步稀释至10-3,取上清液(125 μL)接种到测试板的每个孔中,将接种好的测试板放至25℃下培养,每隔12 h在波长为590 nm的Biolog读数器上读数,实验持续12 d[27]。
1.4 数据处理
根据公式(1)计算测试板孔中溶液吸光值平均变化率(average well color development, AWCD);用OD590>0.15的微孔数作为相应样品土壤Richness指数(用于评估物种的丰富度)[28];根据公式(2)计算Shannon指数H(用于评估物种的丰富度);根据公式(3)计算McIntosh指数U(用于评估群落物种均匀度)[2, 29]。
AWCD =∑ (C-R)/n
(1)
H=-∑Pi(lnPi)
(2)
(3)
式中,C为每个有碳源孔的吸光值,R为对照孔的吸光值,n为碳源孔数,GN板n值为95。Pi为第i孔的相对吸光值与所有反应孔相对吸光值总和的比值,即:Pi=(C-R)/∑(C-R)。
数据处理、相关分析采用Excel 2003软件进行,方差分析、主成分分析和多重比较采用DPS 3.11(Data Processing System for Practical Statistics)软件进行,多重比较采用LSD法(P<0.05);利用CANOCO软件包进行CCA分析。
2 结果与分析
2.1 板孔平均颜色变化率
平均颜色变化率(AWCD)是表示土壤微生物群落利用单一碳源能力的一个重要指标,它反映了土壤微生物活性和群落生理功能多样性[2]。各处理根际土壤AWCD在早稻和晚稻成熟期的变化如图1中所示,各处理根际土壤AWCD整体呈增加的变化趋势。随着培养时间的延长,各处理根际土壤AWCD不断增加;一直持续到第108 h以后,缓慢上升。
在培养时间结束后(276 h),各处理根际土壤AWCD测定结果表明,不同冬季覆盖作物秸秆还田处理对AWCD具有明显的影响(图1)。早稻成熟期,各冬季覆盖作物秸秆还田处理的AWCD均高于冬闲-双季稻(CK)处理,以马铃薯-双季稻(Po)处理均为最高,均显著高于黑麦草-双季稻(Ry)、油菜-双季稻(Ra)和CK处理(P<0.05);其次是紫云英-双季稻(Mv)处理,均显著高于Ra和CK处理(P<0.05)。晚稻成熟期,Po、Ra和Mv处理的AWCD均显著高于CK处理(P<0.05),以Po处理均为最高,均显著高于Ry、Mv和CK处理(P<0.05);其次是Ra处理,均显著高于Ry和CK处理(P<0.05);Mv处理的AWCD均高于Ry处理,但两者间均无显著性差异(P>0.05)。
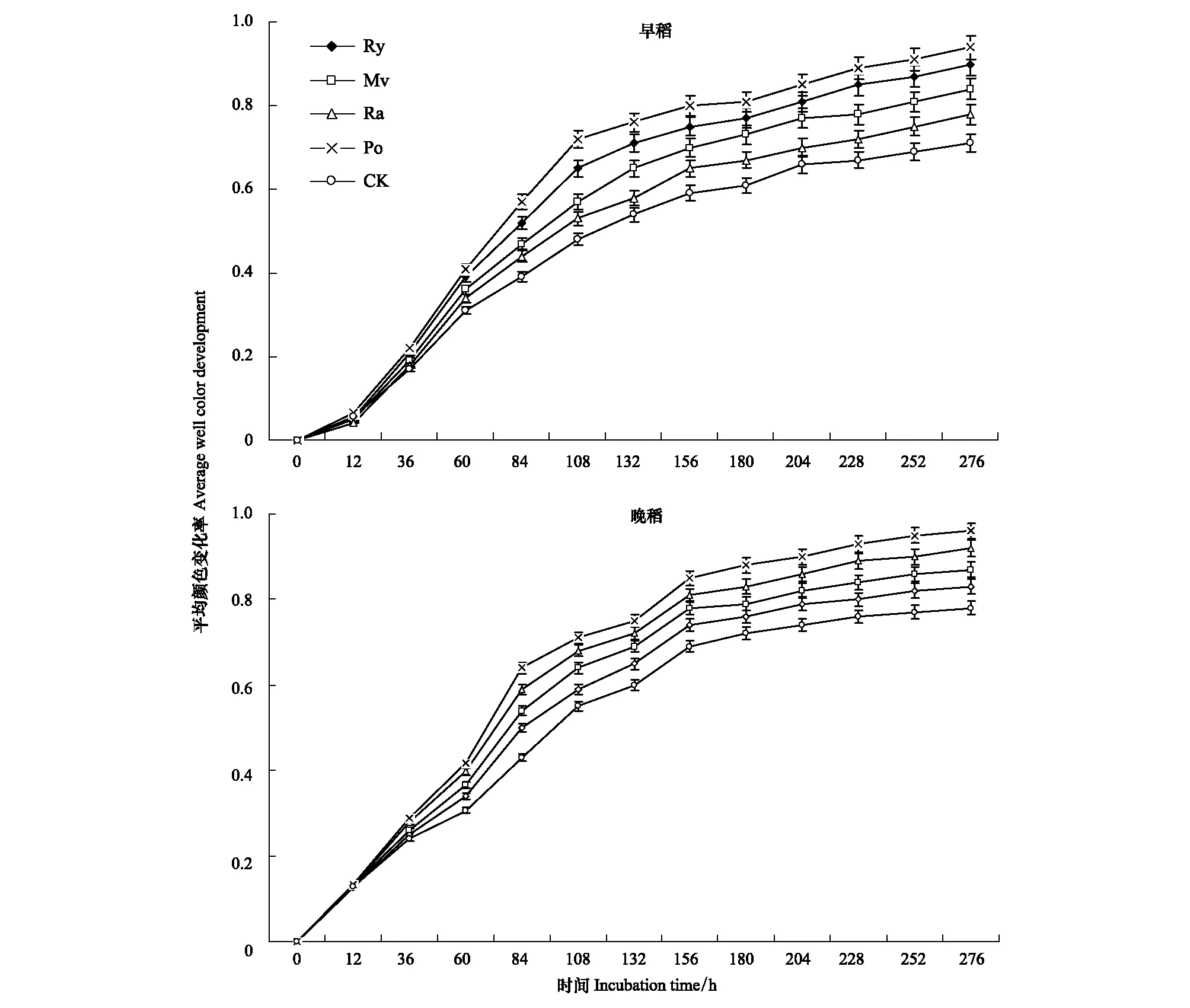
图1 早稻和晚稻成熟期不同冬季覆盖作物秸秆还田处理水稻根际土壤微生物吸光值AWCDFig.1 AWCD changes with incubation progress during early and late rice maturity stages with different straw recycling of winter covering cropsRy:黑麦草-双季稻ryegrass-double cropping rice;Mv:紫云英-双季稻 Chinese milk vetch-double cropping rice;Ra:油菜-双季稻 rape-double cropping rice;Po:马铃薯-双季稻 potato-double cropping rice;CK:冬闲-双季稻 fallow-double cropping rice。结果表示为平均值±标准误 Results stand for means ± standard errors
2.2 土壤微生物群落代谢多样性变化
水稻成熟期的根际土壤Richness指数、Shannon指数和McIntosh指数可反映土壤微生物物种的丰富度、群落物种的均匀度。表2中表明,各冬季覆盖作物秸秆还田处理提高了水稻根际土壤的Richness指数、Shannon指数和McIntosh指数。早稻和晚稻成熟期,Po处理Richness指数和Shannon指数均显著高于CK(P<0.05),其大小顺序分别表现为Po>Ry>Mv>Ra>CK和Po>Ra>Mv>Ry>CK。Po处理的McIntosh指数均为最高,表明马铃薯秸秆还田处理下根际土壤的物种均匀度最高,其次为Mv处理,而冬闲对照处理为最低。试验结果表明,各处理间水稻根际土壤的均匀度与群落丰富度间存在一定的差异。
表2水稻成熟期不同冬季覆盖作物秸秆还田处理根际土壤微生物群落多样性指数值
Table2Geneticdiversityindicesofrhizosphericsoilmicrobialcommunitiesfordifferentstrawrecyclingofwintercoveringcropstreatmentsatearlyandlatericematuritystages

水稻Rice处理TreatmentsRichness指数 Richness indexShannon指数 Shannon indexMcIntosh指数 McIntosh index早稻Early riceRy16.43±0.48ab2.64±0.07a5.68±0.14bcMv16.05±0.45ab2.52±0.07ab6.06±0.17abRa15.68±0.46ab2.47±0.08bc5.18±0.18cPo16.77±0.47a2.73±0.07a6.25±0.16aCK15.04±0.43b2.34±0.06c4.63±0.13d晚稻Late riceRy13.87±0.43ab2.45±0.08bc5.32±0.18cMv14.16±0.42ab2.57±0.07ab5.72±0.17bcRa14.48±0.41ab2.69±0.07a6.14±0.16abPo14.92±0.40a2.78±0.07a6.38±0.15aCK13.28±0.38b2.33±0.06c4.74±0.14d
Ry:黑麦草-双季稻ryegrass-double cropping rice;Mv:紫云英-双季稻 Chinese milk vetch-double cropping rice;Ra:油菜-双季稻 rape-double cropping rice;Po:马铃薯-双季稻 potato-double cropping rice;CK:冬闲-双季稻 fallow-double cropping rice。结果表示为平均值±标准误 Results stand for means ± standard errors。同列数据后不同字母表示差异达到5%的显著水平 Means in each column with different letters are significantly different at theP< 0.05 level
2.3 水稻根际土壤微生物群落代谢功能主成分分析
根据前人的研究结果,所提取的成分一般要求其累计方差贡献率达到85%的原则[30],在本试验中共提取了8个主成分,累计贡献率分别达86.22%和87.35%。其中,早稻成熟期第1主成分(PC1)的方差贡献率为33.26%,第2主成分(PC2)为16.57%,第3—8主成分贡献率均较小,分别为8.85%、7.75%、6.28%、5.38%、4.28%和3.85%;晚稻成熟期,PC1的方差贡献率为35.57%,PC2为17.58%,第3—8主成分贡献率均较小,分别为8.63%、7.15%、6.26%、4.57%、4.03%和3.56%,因此本文只对前2个主成分进行分析(图2)。
分析结果表明,不同冬季覆盖作物秸秆还田处理在PC轴上存在明显的差异,在PC1轴上各处理分布分散,CK处理主要分布在负方向,而Po、Ra和Ry处理主要分布在PC1轴正方向;PC2轴上CK处理分布在负方向,Po处理分布在正方向,其中Ry处理在PC2正、负轴上均有分布。
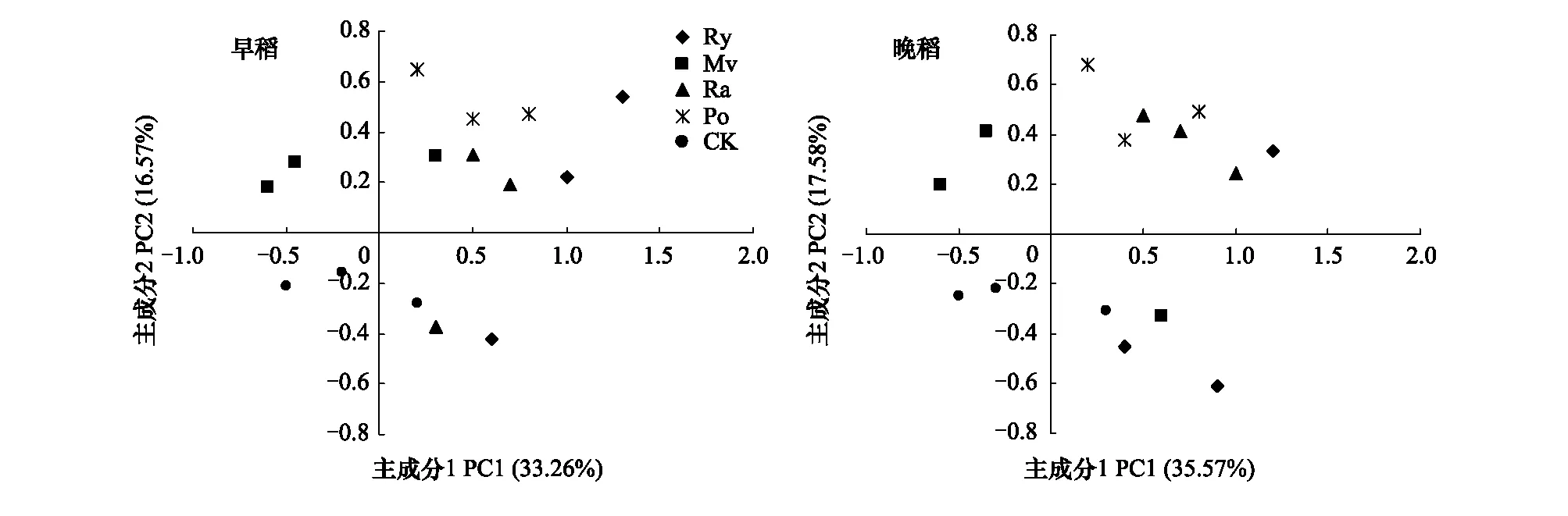
图2 水稻成熟期不同冬季覆盖作物秸秆还田处理根际土壤微生物碳源利用类型主成分分析Fig.2 Principal components analysis for carbon utilization of rhizospheric soil microbial communities in different straw recycling of winter covering crops treatments at early and late rice maturity stages
按与PC1和PC2相关系数的大小,本研究选择对位于前12个碳源进行分析,分析结果显示,对PC1和PC2影响作用较大的碳源物质主要氨基酸和糖类物质(表3)。在PC1中,碳源为氨基酸类物质所占比例达58.3%;而在PC2中,碳源为糖类物质所占比例达83.3%;因此,区分各个不同冬季覆盖作物秸秆还田处理间碳源的主要物质为氨基酸和糖类物质。
表3土壤中与PC1和PC2相关显著的主要培养基
Table3MainsubstrateswithhighcorrelationcoefficientsforPC1andPC2PCAofdiversitypatternsfordifferentstrawrecyclingofwintercoveringcropstreatments

主成分1 PC1主成分2 PC2培养基类型 Substratesr培养基类型 Substratesr培养基类型 Substratesr培养基类型 SubstratesrD-甘露醇(糖类)D-Mannitol0.875L-脯氨酸(氨基酸类)L-Prolin0.809L-天门酰氨酸(氨基酸类)L-Asparagine0.874L-阿拉伯糖(糖类)L-Arabinos0.802D-葡萄糖胺酸(羧酸类)D-Gjucosam inicacid0.856D-丙氨酸(氨基酸类)D-Alanine0.789麦芽糖(糖类)Maltose0.867D-果糖(糖类)D-Fructose0.793L-丝氨酸(氨基酸类)L-Serine0.841L-天门冬氨酸(氨基酸类)L-Asparticaci0.781β-甲基-D-葡萄糖苷(糖类)L-Methyl-D-glucoside0.856D-甘露醇(糖类)D-Mannitol0.745L-焦谷氨酸(氨基酸类)L-Pyroglutamic acid0.833甲基丙酮酸(酯类)Pyruvic acid methylester0.753D-纤维糖(糖类)D-Cellobisoe0.836N-乙酰-D-葡萄糖胺(糖类)N-Acetyl-D-glucosamine0.734L-亮氨酸(氨基酸类)L-Leucine0.825D-葡萄糖酸(羧酸类)D-Gluconic acid0.7456-磷酸葡萄糖(糖类)Glucose-6-phosphate0.823D-木糖醇(糖类)D-Xylose0.713α-D-葡萄糖(糖类)α-D-Glucose0.812γ-氨基丁酸(氨基酸类)γ-Aminobutyric acid0.7261-磷酸葡萄糖(糖类)Glucose-1-phosphate0.815丙二酸(羧酸类)Malonic acid0.705
本试验选择对不同冬季覆盖作物秸秆还田处理具体利用的前6种碳源进行分析,不同冬季覆盖作物秸秆还田处理根际土壤微生物利用的碳源物质代码如表4中所示。稻田土壤化学性质变化与根际土壤微生物碳源利用物质之间的相互关系分析结果表明,从稻田土壤微生物碳源利用组成在双轴空间上的分布来看,糖类和羧酸类物质主要分布在轴1的右边,氨基酸类、酯类和部分糖类物质主要分布在轴1的左边。轴1右边与土壤pH、有机质、全氮、碱解氮含量均表现出正相关,糖类和羧酸类物质主要受土壤pH、有机质、全氮、碱解氮含量的影响;土壤有效磷和速效钾含量均是影响氨基酸类和部分糖类物质的主要环境变量(图3)。
表4CCA分析中各处理微生物利用的主要碳源代码
Table4Codesofmaincarbonsubstratesutilizedbyrhizosphericsoilmicrobialcommunitiesunderdifferentstrawrecyclingofwintercoveringcropstreatmentsforcanonicalcorrespondenceanalysis

编号 Code碳源 Carbon substrates编号 Code碳源 Carbon substratesA1D-甘露糖(糖类) D-MannoseB1D-纤维糖(糖类) D-CellobisoeA2L-焦谷氨酸(氨基酸类) L-Pyroglutamic acidB2L-阿拉伯糖(糖类) L-ArabinoseA3D-半乳糖(糖类) D-GalactoseB3N-乙酰-D-葡萄糖胺(糖类) N-Acetyl-D-glucosamineA4D-葡萄糖酸(羧酸类) D-Gluconic acidB4D-甘露醇(糖类) D-MannitolA5L-丝氨酸(氨基酸类) L-SerineB51-磷酸葡萄糖(糖类) Glucose-1-phosphateA6D-丙氨酸(氨基酸类) D-AlanineB6L-天门酰氨酸(氨基酸类)L-AsparagineA7甲基丙酮酸(酯类) Pyruvic acid methylesterB7β-甲基-D-葡萄糖苷(糖类) L-Methyl-D-glucosideA8L-亮氨酸(氨基酸类) L-LeucineB8D-果糖(糖类) D-FructoseA9D-葡萄糖胺酸(羧酸类)D-Gjucosam inicacidB9N-乙酰基-D-半乳糖胺(糖类) N-Acetyl-D-galactosamineA10L-天门冬氨酸(氨基酸类) L-AsparticaciB10D-木糖醇(糖类) D-XyloseA11L-脯氨酸(氨基酸类) L-ProlinB116-磷酸葡萄糖(糖类) Glucose-6-phosphateA12α-D-葡萄糖(糖类) α-D-GlucoseB12丙二酸(羧酸类) Malonic acidA13γ-氨基丁酸(氨基酸类) γ-Aminobutyric acidB13麦芽糖(糖类) Maltose
3 讨论
前人研究结果表明,土壤碳源平均颜色变化率(AWCD)大小可用来表示土壤微生物对碳源利用能力和代谢活性的大小,作为土壤微生物活性的有效指标[31]。周文新等[14]研究认为,稻草还田有利于改善土壤微生物群落结构和功能,提高土壤微生物群落代谢能力(AWCD)和功能多样性(丰富度与多样性指数)。在本研究中,不同冬季覆盖作物秸秆还田措施对稻田根际土壤微生物群落碳源利用能力具有明显的影响。与冬闲-双季稻对照相比,各冬季覆盖作物秸秆还田措施均增加了AWCD,这表明施入外源冬季覆盖作物秸秆措施均有利于维持根际土壤微生物的碳源利用能力,其原因可能是由于外源有机物(冬季覆盖作物秸秆)的施用,不同的有机物中含有丰富的碳含量和营养物质,还田后能为土壤微生物的活动提供丰富的碳源和物质来源,增加了土壤微生物的活性[11]。在各个冬季覆盖作物秸秆还田处理中,均以马铃薯秸秆还田措施增加AWCD效果为最佳,这可能是由于马铃薯茎叶、稻草秸秆还田配施化肥后N、P、K养分均衡且C/N适宜,秸秆中养分释放较快,促进了微生物的分解[32];另一方面,马铃薯秸秆还田处理可能改变了根际土壤微生物的优势种群,增加了其对所测试碳源的利用率;同时,马铃薯秸秆还田量、秸秆中碳含量和碳还田量也可能是影响与土壤碳循环微生物活动的重要因素[33]。早稻成熟期,黑麦草和紫云英秸秆还田措施也增加了AWCD,这可能是黑麦草和紫云英秸秆在早稻生育期较易进行分解、释放养分,促进了土壤微生物的活性[23],从而增强了微生物对碳源的利用率。晚稻成熟期,油菜秸秆还田措施也增加了AWCD,其原因可能是油菜还田秸秆在早稻生育期的分解进程较缓慢、晚稻生育期仍有部分秸秆进行分解、释放养分[22- 23],从而促进了土壤微生物的活动,增加了其AWCD。
前人研究结果表明,物种丰富度(Richness指数、Shannon指数)和均匀度指数(McIntosh指数)是表征土壤微生物群落多样性的常用指标[2]。在本研究中,对各处理土壤Richness、Shannon和McIntosh指数分析结果显示,不同冬季覆盖作物秸秆还田措施对稻田根际土壤微生物群落多样性有显著的影响,各个秸秆还田处理中根际土壤微生物物种的数量、各个物种的均匀度均有差异。在不同冬季覆盖作物秸秆还田措施条件下,马铃薯秸秆还田措施最有利于提高根际土壤微生物群落物种数量、各物种的均匀度,这可能是由于马铃薯茎叶、稻草秸秆还田配施化肥后N、P、K养分均衡且C/N适宜有利于根际土壤微生物的生长;且所释放的养分有利于培肥土壤,能为土壤微生物提供较多的能源与养分,促进根际土壤微生物大量繁殖[34],加快微生物的新陈代谢;同时,马铃薯茎叶、稻草秸秆中均含有丰富的碳含量[33],该种还田措施下增加了土壤中C的还田量,增强了与碳利用相关土壤微生物活性。因此,马铃薯秸秆还田措施能显著提高根际土壤微生物活性,增强了碳源利用能力(图1),而其群落多样性也得到提高。与冬闲-双季稻对照相比,黑麦草和紫云英秸秆还田措施增加了早稻根际土壤微生物的物种丰富度,油菜和紫云英秸秆还田措施增加了晚稻根际土壤微生物的物种丰富度和均匀度指数,这可能是黑麦草和紫云英秸秆还田后在早稻生育期较易进行分解、释放养分[35- 36],为土壤微生物的活动提供了物质来源,增加微生物物种的数量;而油菜和紫云英秸秆在晚稻生育期仍有部分秸秆进行分解、释放养分[22- 23],也促进了土壤微生物的活动,增强了土壤微生物种群的生长代谢,从而增加了根际土壤微生物的物种丰富度和群落均匀度。在早稻和晚稻季,各冬季覆盖作物秸秆还田处理间根际土壤的物种丰富度和均匀度指数大小顺序均存在一定的差异,这可能与冬季覆盖作物秸秆类型、秸秆还田量[22- 23]、秸秆中碳含量[19, 33]、碳还田量[19- 20, 33]、还田后在土壤中的分解速率[35- 36]、早稻和晚稻生育期外界气候条件[21]不同密切相关。
前人研究认为,主成分分析表明在不同措施下土壤微生物碳源利用能力的差异水平,各样品在主成分轴上分布与微生物对碳源底物利用能力的关系密切[12]。Garland等[2]研究结果表明,各试验样本在PC轴坐标上的差异与在该PC轴上碳源利用能力之间是相互对应的。在本试验的主成分分析中,对前2个主成分进行了分析,结果表明不同冬季覆盖作物秸秆还田措施下根际土壤微生物的碳源利用能力存在显著差异(图2)。其原因是一方面,土壤有机物质含量和组成变化及土壤化学性质变化是影响不同冬季覆盖作物秸秆还田措施下土壤微生物碳源代谢特征的重要原因之一,冬季覆盖作物秸秆中含有大量碳水化合物以及氮、磷、钾等营养元素,还田后影响稻田土壤有机物质含量和组成、部分土壤养分含量[20]、土壤化学性质与根际土壤微生物碳源利用物质之间相关性(图3),使土壤微生物群落发生很大变化[37]。另一方面,土壤团聚体是微生物生长的重要微环境,土壤微生物与土壤团聚体数量及质量之间存在密切的关系,不同冬季覆盖作物秸秆还田后对土壤团聚体具有明显的影响[19],影响到土壤微生物群落特征,使土壤微生物碳源代谢类型发生改变。在根际土壤微生物利用碳源方面,周文新等[14]研究认为,稻草还田处理所利用的碳源主要包括β-甲基—D-糖化物、甲基丙酮酸D-木糖醇等9种物质。在本试验冬季覆盖作物-双季稻种植模式条件下,采取不同冬季覆盖作物秸秆还田措施双季稻田根际土壤微生物利用的主要碳源为氨基酸类和糖类物质(表3),这可能是因为土壤中加入外源有机物质后,显著提高了氨基酸和单糖等物质的含量,土壤微生物可以利用的碳源趋于稳定[12],促进偏好氨基酸类、单糖糖类物质为碳源的微生物群落的发育;但不同的冬季覆盖作物秸秆还田措施处理间根际土壤微生物所利用的氨基酸和糖类等物质存在明显的差异,其原因为不同类型秸秆还田后对土壤化学性质具有明显的影响[20],影响与根际土壤微生物碳源利用物质之间相关性(图3),对各处理根际土壤微生物所利用物质造成显著的差异,各处理间根际土壤微生物所利用的物质类型可作为划分各处理间对根际土壤微生物影响的依据[38- 39]。
4 结论
与冬闲-双季稻处理相比,各冬季覆盖作物秸秆还田处理提高了双季稻田根际土壤微生物对碳源的利用程度(AWCD)。冬季覆盖作物秸秆还田处理均有利于维持双季稻田根际土壤微生物群落多样性,特别是马铃薯秸秆还田处理能提高稻田根际土壤微生物物种的丰富度和群落物种的均匀度,冬闲对照处理降低了稻田根际土壤微生物群落均匀度。土壤微生物碳源利用主成分分析结果表明,各冬季覆盖作物秸秆还田处理根际土壤微生物利用的主要碳源为氨基酸类和糖类物质,不同冬季覆盖作物秸秆还田处理间稻田根际土壤微生物群落在碳源利用类型上存在明显的差异,冬季覆盖作物秸秆还田措施增加了稻田根际土壤微生物群落在碳源利用上的变异程度。运用典范对应分析(CCA)方法对稻田土壤化学性质变化与根际土壤微生物碳源利用物质之间的关系分析结果表明,不同冬季覆盖作物秸秆还田处理根际土壤微生物利用的主要碳源糖类和羧酸类物质主要受土壤pH、有机质、全氮、碱解氮含量的影响,土壤有效磷和速效钾含量均是影响氨基酸类和部分糖类物质的主要环境变量。
猜你喜欢
今日农业(2021年21期)2022-01-12
中国土壤与肥料(2021年5期)2021-12-02
天然产物研究与开发(2018年3期)2018-05-07
中国蔬菜(2016年8期)2017-01-15
天然产物研究与开发(2016年1期)2016-06-05
中国民族医药杂志(2016年7期)2016-05-09
上海农业学报(2016年5期)2016-02-10
湖南农业(2015年5期)2015-02-26
植物营养与肥料学报(2012年1期)2012-10-26
植物营养与肥料学报(2011年2期)2011-10-26
