
太白山栎属树种气孔特征沿海拔梯度的变化规律
2018-11-02 06:30温婧雯陈昊轩滕一平张硕新王瑞丽
生态学报 2018年18期
温婧雯,陈昊轩,滕一平,张硕新,2,王瑞丽,*
1 西北农林科技大学林学院,杨凌 712100 2 陕西秦岭森林生态系统国家野外科学观测研究站,杨凌 712100
气孔是植物体内水气与外界环境交换的主要通道,并且控制植物的光合、呼吸和蒸腾作用[1]。植物通过气孔开闭对短时间的环境变化快速做出反应,因此气孔形态特征是植物在进化生存的过程中对外界环境长期适应的结果[2- 3]。单位叶表面积上气孔数量(即气孔密度)与气孔大小共同决定了最大可能气孔导度,最大气孔导度直接影响最大同化速率的大小[4- 5]。气孔密度与气孔大小调节气孔的发育与运动使得这两个相互矛盾的需求达到最优化[2],小的气孔具有短的反应时间,往往与高气孔密度相连,使叶片在有利条件下快速获得较高的气孔导度[6- 7]。探究气孔性状与环境的关系对了解植物进化规律及其应对并影响全球气候变化具有重要意义。
海拔作为一种重要的地形因子,在较小的范围内就可以形成环境与性状的强烈变化,这为研究植物对环境变化的响应提供了理想的场所[8- 9]。长期以来,关于气孔性状对海拔梯度的响应展开大量研究并取得一些结果。随海拔升高,气孔密度显著升高[10]、或显著下降[11]、又或无显著变化[12]等。Qiang等[13]研究表明,青海云杉(Piceacrassifolia)气孔密度随海拔先升高后降低,辽东栎(Quercuswutaishansea)则相反[14]。Vitousek等[15]对多型铁心木(Metrosiderospolymorpha)的研究发现气孔导度随海拔的升高而降低,与Friend等[16]研究结果相反。气孔大小(气孔长度、宽度或二者的乘积)随海拔变化的形式也各不相同[12,14,17]。性状与环境的关系能够反映出不同植物在自然条件下的适应策略[18]。此外,各性状之间不是独立存在的,植物常常通过性状组合来适应不断变化的环境[4,6]。已有研究证明气孔大小与气孔密度之间存在稳定的负相关关系[4,6,12,14],气孔密度与叶片的水分散失显著正相关,因而较高的气孔密度往往代表了较高的气孔导度[6- 7,19],这些研究都表明气孔性状之间存在一定的关联性。然而,以往关于气孔性状的研究多侧重于所有植物的普遍规律或单一植物的试验研究[4,10,14,17],而对亲缘关系相近的不同植物间的差异比较研究较少。因此,本文拟选择同一属的不同植物为研究对象,探讨亲缘植物的气孔性状随海拔梯度的变化规律,这有助于理解全球气候变化背景下植物的生理适应特征。
秦岭是中国地理南北分界线和气候、生态分水岭。太白山是秦岭主峰和最高峰,位于秦岭山脊线处,高差大,坡度陡,地貌特征复杂,是对全球变化响应敏感的地区之一,也是研究植物沿海拔梯度变化的理想地点[20-21]。壳斗科(Fagaceae)栎属(Quercus)是森林树种中分布最广的类群,生态适应幅度大,许多种类是群落中的优势种或建群种[21-22],对环境具有高度适应能力和在复杂生境下的自我调控能力,其叶片对气候变化非常敏感,探讨栎属树种对环境变化的生态适应策略具有重要意义。因此本文以太白山4种栎属树种叶片气孔为研究对象,在野外调查的基础上,选择气孔密度、气孔长度、气孔宽度以及潜在气孔导度指数等气孔性状指标,旨在研究:(1)太白山海拔变化如何影响4种栎属树种气孔性状? (2)这些影响在不同树种间是否有差异?
1 研究区概况
研究区位于秦岭主峰太白山自然保护区(107°41′23″—107°54′40″E,33°49′31″—34°08′11″N),海拔1100—2300 m之间。该区域属于典型的内陆季风气候区,夏季炎热多雨,冬季寒冷干燥,其北坡年降水量为694.2 mm,年均温13℃[23]。在海拔800—2300 m之间主要为落叶阔叶林带,其中在海拔800—1400 m间为栓皮栎(Q.variabilis)林亚带,土壤属典型褐土;海拔1000—1900 m间为锐齿栎(Q.alienavar.acuteserrata)林亚带,土壤以普通棕壤为主,间有弱灰化棕壤;在海拔1900—2300 m间为辽东栎(Q.wutaishansea)林亚带,土壤为普通棕壤与弱灰化棕壤,间有生草棕壤[21]。研究区内栎属树种分布垂直地带性明显,自下而上有栓皮栎、槲栎(Q.aliena)、锐齿栎、辽东栎。
2 研究方法
2.1 野外取样

图1 太白山栎属树种海拔取样图 Fig.1 Sampling diagram of species of Quercus in Taibai Mountain ●为取样树木
2016年7月以100 m海拔间隔,采集太白山北坡1100—2300 m海拔梯度的栎属树种样品,选取地形、地貌、坡度和坡向等基本一致的标准样地,按“ S ”型方法选择健康样木进行取样(图1)。在每个海拔梯度上,每个树种选择4个树高、胸径均匀的个体,每个个体选取位于树冠外围、能最大程度的接收阳光照射、完全成熟、无病虫害损坏的叶片30片。同一海拔的混交树种样品分开,装入10号自封袋内,带回实验室放入冰箱。同时,在距离所选样木1 m处,利用土钻对0—10 cm的土层进行取样,去除根系和石砾后混合均匀。待土壤样品风干后全部过2 mm筛,利用元素分析仪测定土壤有机碳、全氮和全磷含量,同时测定土壤的湿度、温度和pH值[24]。
2.2 室内测定
气孔密度(Stomatal density,SD)的测定采用印迹法,详见王瑞丽等[24]。具体步骤为:每一个树种个体随机选择3个叶片,首先使用透明指甲油涂在叶片下表皮(大约1 cm2,注意避开主脉和大的次级脉),静置待指甲油干透后用镊子撕下,制成临时切片。然后将切片置于显微镜下,并利用配置的摄像装置进行观测,在40倍物镜、10倍目镜条件下拍照,每一样品上随机选择3—5个视野记录气孔数目,根据视野内的气孔数量计算SD。同时在每个视野图片中随机选择3个气孔,利用配套的分析软件测定气孔长度(Stomatal length,SL)、气孔宽度(Stomatal width,SW),本文以气孔器保卫细胞的长轴长度(μm)和短轴长度(μm)代表气孔大小。
气孔可塑性指数按Valladares[25]的方法计算,即可塑性指数(Phenotypic plasticity index,PI)=(最大值-最小值)/最大值。
由于气孔密度和气孔大小共同决定最大气孔导度(gmax)[4- 5,26],采用Holland等[12]方法计算潜在气孔导度指数 (Potential conductance index, PCI)。
PCI=SL2×SD×10-4
2.3 气象数据的获取
由于太白山地形复杂,野外气候考察往往只是短期的行为,结果不具有代表性,因此山地研究中所需要的气候资料大多是通过山区周边地区的气候资料推算得到[27]。
本研究将唐志尧等[23]推算方法作为气温(Mean annual temperature,MAT)、相对湿度(Relative humidity,RH)数据的推算依据,以傅抱璞等[28]得出的推算公式作为降水(Mean annual precipitation,MAP)数据的推算依据,推算出每个海拔高度的气温、相对湿度和降水量。
MAT=-0.0049ALT+17.9,(R2=0.99,P<0.001)
RH=0.4 ×10-6(ALT)2-0.0153ALT +83.7(R2=0.95,P<0.001)
MAP(Pz)=Ph0+a[ (2H-Z)·Z-(2H-h0)·h0]
式中,ALT为海拔高度(m),Pz为某一海拔高度降水量;Ph0为最大降水高度(H)以下某一参考高度(h0)的降水量;Z为海拔高度;a为区域参数,秦岭北坡a=4.795×10-5[29]。
2.4 数据分析
首先,对所有气孔性状进行以10为底的对数转换,使数据满足标准化正态分布,随后计算气孔形态特征间Pearson相关系数;其次,针对存在极显著的性状关系采用标准化主轴估计(Standardized Major Axis Estimation, SMA) 方法[30],来分析气孔性状关系是否在不同树种间存在显著差异,即利用方程y=bxa表示不同气孔性状间的关系,线性转化为ln(y)=ln(b)+aln(x),x、y表示两个不同性状,b代表性状关系的截距,a表示相关性的斜率。该分析由软件(S) MATR Version 2.0[31]来计算完成。最后,对气孔性状与环境因子进行相关性及一般线性模型(GLM)分析。实验所用数据采用SPSS 20.0软件进行处理,相关图形用SigmaPlot 10.0以及OriginPro 8软件绘制。
3 结果
3.1 太白山栎属树种气孔性状概况
栎属树种气孔性状概况见表1。叶片气孔性状中,SD、SL和SW的均值分别为678.27个/mm2、21.08 μm、15.18 μm,变化范围为404.00—1072.00个/mm2、17.69—26.99 μm、12.01—27.38 μm,其中,SD的变异程度(CV=0.25)大于SL(CV=0.09)和SW(CV=0.16)。PCI的变化范围为17.56%—45.05%,平均值为29.67%,变异系数为0.22。由表2可见,锐齿栎和栓皮栎PCI的可塑性指数较大,分别为0.446、0.392;锐齿栎SD的可塑性指数最大,为0.397。
对气孔性状进行差异显著性分析表明(图2):栓皮栎的SL、SW、PCI与槲栎、锐齿栎和辽东栎之间差异显著(P<0.05)。辽东栎与锐齿栎的SW、SL之间差异不显著(P>0.05),SD与PCI差异显著(P<0.05)。槲栎的SD、PCI与辽东栎差异显著(P<0.05)。其中,栓皮栎的SL最大,辽东栎次之;槲栎的SW最小,栓皮栎最大。锐齿栎和槲栎的SD与PCI较大且无显著差别,栓皮栎SD最小,辽东栎的PCI最小。
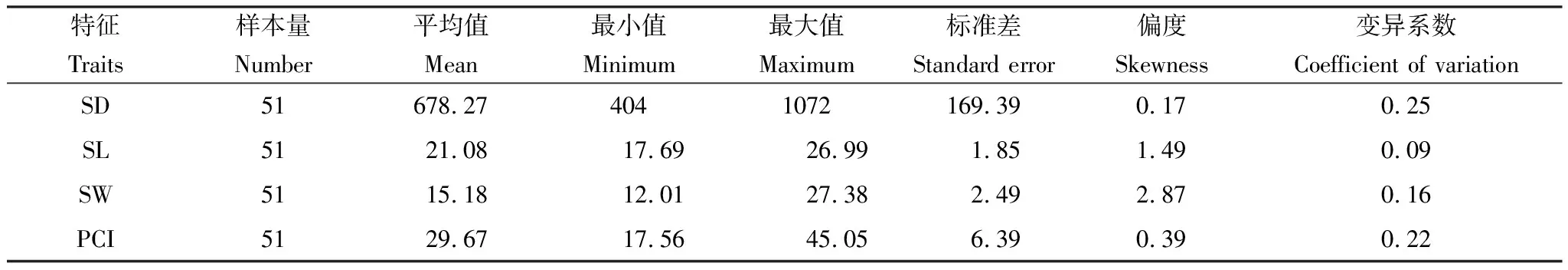
表1 气孔性状特征的数据描述
SD:气孔密度 stomatal density;SL:气孔长度 stomatal length;SW:气孔宽度 stomatal width;PCI 潜在气孔导度指数 potential conductance index

表2 4种栎属树种气孔性状的可塑性指数
栓皮栎:Quercusvariabilis;槲栎:Q.aliena;锐齿栎:Q.alienavar.acuteserrata;辽东栎:Q.wutaishansea
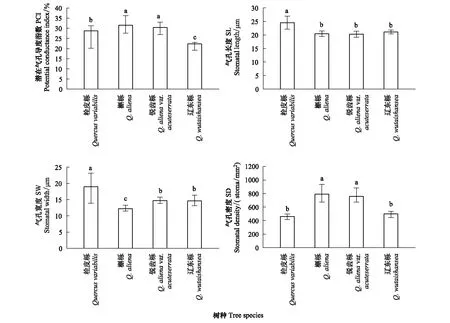
图2 太白山4种栎属树种气孔特征性状值的变化趋势 Fig.2 Trends of stomatal traits in four species of Quercus from Taibai MountainSD:气孔密度 stomatal density;SL:气孔长度 stomatal length;SW:气孔宽度 stomatal width;PCI 潜在气孔导度指数 potential conductance index
3.2 气孔性状间的关系
如表3所示,SD与其他气孔性状的关系,均达到极显著水平(P<0.01)。其中,SD与SL和SW负相关,与PCI正相关。SL与SW存在极显著的正相关(P<0.01)。
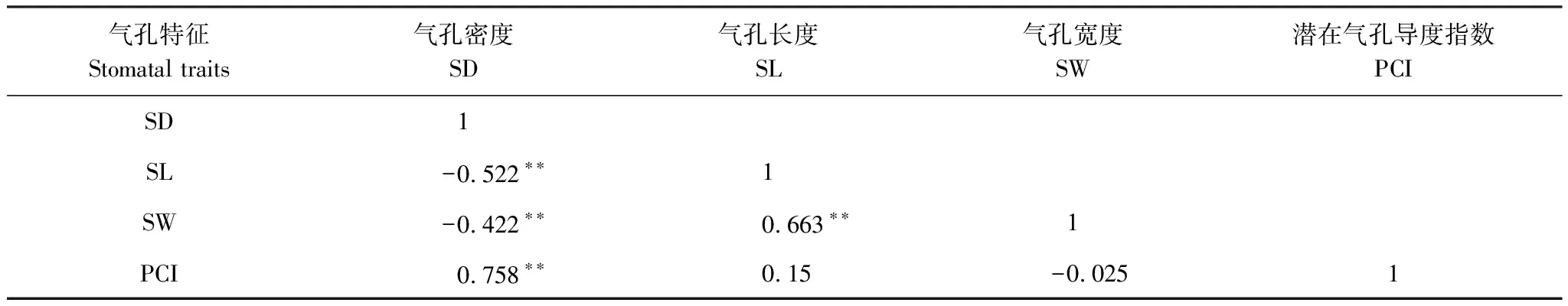
表3 叶片气孔特征间的Pearson相关系数
**,P<0.01
进一步分析表明,除栓皮栎外,SD与PCI的正相关关系在其他树种间均达到显著水平(图3,P<0.05)。SW与SL之间的正相关关系只在栓皮栎和锐齿栎之间达到显著水平(图3,P<0.05)。
SD与SL之间存在稳定的负相关关系,这种负相关关系在不同树种之间均存在(图3)。不同树种间斜率差异不显著(SMA共同斜率为-2.076,P>0.05),但截距间存在显著差异(P<0.05)。其中,辽东栎处的截距最大,栓皮栎处的截距最小。各个方程参数见表4。

图3 不同栎属树种气孔密度与潜在气孔导度指数、气孔宽度与气孔长度、气孔密度与气孔长度的标准化主轴回归关系Fig.3 SMA regression of stomatal density and potential conductance index, stomatal width and stomatal length,stomatal density and stomatal length for different Quercus species只列出显著的回归关系(P<0.05);*, P<0.05;**, P<0.01

指标物种组样本量相关系数显著水平斜率95%置信区间IndexGroupNumberR2PSlope95% CIsSL与SWH190.0030.836L360.0530.175R810.0770.0121.3171(1.0639,1.6306)S180.779<0.00010.8135(0.6358,1.0408)SD与PCIH190.4470.0021.045(0.720,1.516)L360.605<0.00011.191(0.958,1.480)R810.4630.0421.297(1.101,1.527)S180.2340.052SD与SLH190.1360.021-2.407(-3.811,-1.521)L360.7380.003-2.672(-3.759,-1.899)R810.2290.018-2.088(-2.602,-1.676)S180.220.049-1.03(-1.620,-0.655)
S:栓皮栎Quercusvariabilis;H:槲栎Q.aliena;R:锐齿栎Q.alienavar.acuteserrata;L:辽东栎Q.wutaishansea
3.3 气孔性状沿海拔梯度的变化
叶片气孔密度、气孔长度和潜在气孔导度指数等性状沿海拔梯度呈曲线变化形式(图4)。在海拔约1600 m处,气孔密度和潜在气孔导度指数的平均值达到最大(分别为754.36个/mm2和32.74%),而气孔长度的平均值达到最小(19.70 μm),气孔宽度沿海拔下降缓慢,斜率为-0.0014,具有相对稳定性。
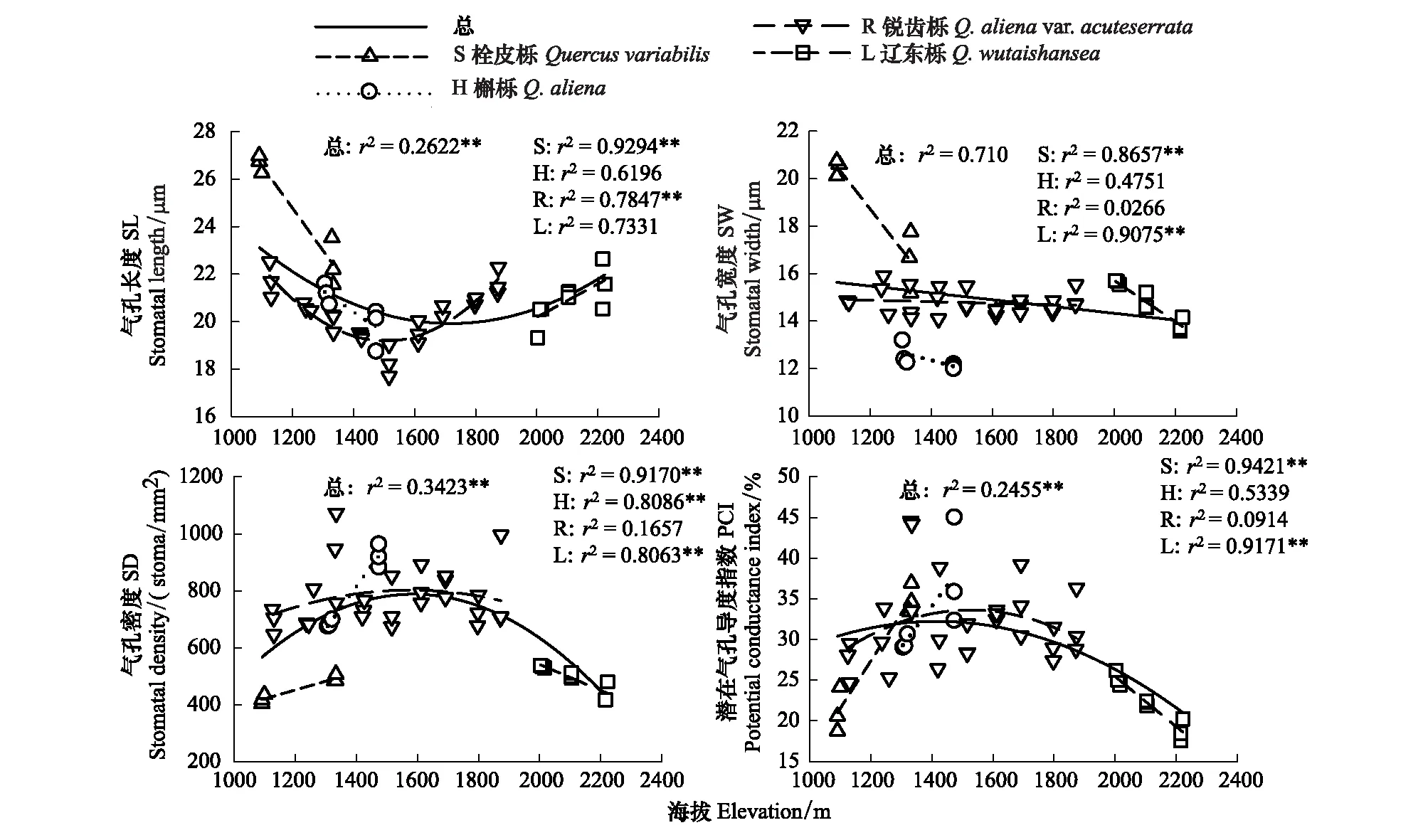
图4 太白山栎属树种气孔性状沿海拔梯度的变化Fig.4 Variation of stomatal characteristics of Quercus trees in Taibai Mountains along the altitudinal gradient*,P<0.05;**,P<0.01
4种树种气孔性状沿海拔变化各不相同。栓皮栎和槲栎的SL、SW随海拔升高下降明显,SD、PCI增加;锐齿栎的SW沿海拔保持相对稳定,变化不明显,SL先降低后升高,在约1600 m处达到最小值,同时SD及PCI达到最大值,之后随海拔升高降低;辽东栎SL逐渐增大,SW降低,同时,SD与PCI沿海拔降低。
3.4 叶片气孔性状与环境因子的关系
3.4.1 气孔性状与土壤因子的关系
气孔性状与土壤因子的相关分析结果如表5所示。土壤湿度对SL存在显著影响(P<0.05),土壤总碳、总磷等因子对气孔性状均无显著性影响(P>0.05)。
3.4.2 气孔性状与气候因子的关系
气孔性状与气候因子的相关分析显示,随着年均降水量的增加,气孔长度、气孔宽度及潜在气孔导度指数均随之下降;同时,潜在气孔导度指数随着温度及湿度的升高均升高,且显著存在(表5)。然而,各环境因子对气孔密度的影响均未达到显著水平(P>0.05)。
进一步分析各环境因子对气孔性状的解释量可看出,气候因子对潜在气孔导度指数影响的大小为:湿度>温度>降水量,解释变异信息的量分别为22.90%、16.70%、4.31%;降水量是影响气孔长度与宽度的主要因子,解释量分别为4.10%和4.60%(表6)。

表5 栎属树种叶片功能性状与环境因子间相关关系
*,P<0.05;**,P<0.01; MAP:mean annual precipitation;MAT:mean annual temperature;RH:relative humidity;TC: soil total carbon;TP:soil total phosphorus;H:soil humidity
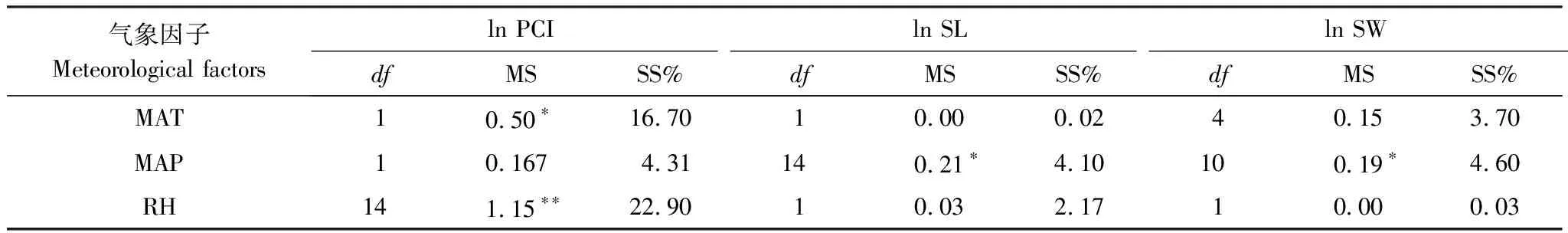
表6 PCI、SL、SW的一般线性模型(GLM)结果
MS,均方 mean squares; SS%,解释平方和百分比ercentage of sum of squares explained;*,P<0.05;**,P<0.01
4 讨论
4.1 气孔性状间的关联性
气孔性状在不同物种间存在较大差异,不同气孔性状间关系紧密,植物往往通过性状组合来适应所在环境的变化[4,6]。
4种栎属树种的气孔密度(SD)与气孔长度(SL)之间存在稳定的负相关关系,该结果与其他对多物种及不同时间地点的研究结果基本一致[4,6,24]。单位面积内气孔数量多,气孔小的叶片能够更好地储存水分,在干旱胁迫时期能够保持稳定的水分供应,这种权衡关系调控了植物气孔形态特征对生长环境的短期可塑性与长期的适应性进化[32]。
SD与植物叶片的水分蒸腾散失显著正相关,因而较高的SD往往代表较高的气孔导度和光合速率[4,6-7,19]。与之类似,本研究中除栓皮栎外,锐齿栎、槲栎及辽东栎的潜在气孔导度指数(PCI)与SD显著相关,且随SD增加呈线性递增。其中锐齿栎的变化斜率最大,说明SD一定时,锐齿栎往往具有更大的气孔导度和碳同化速率。锐齿栎对环境变化比较敏感,因而在太白山北坡,锐齿栎是分布区域最广的优势树种[21,23]。SL与气孔宽度(SW)的相关性只在锐齿栎和栓皮栎中显著存在(P<0.05),与锐齿栎相比,栓皮栎SL与SW的关系更紧密,这可能是由于栓皮栎多分布在低海拔地区(800—1400 m),生存环境比较适宜,因而变异性较小[24]。
本研究结果发现,气孔性状间具有一定的关联性,但这种关联性并非在每个树种内都显著存在(P<0.05,图3),可能的原因有:(1)单一植物的性状值变化较小。生长环境相近的植物性状具有相似性,可能会限制性状之间关系的表达[33]。(2)生存环境不同。植物分布受到许多环境因子的共同影响,每个海拔梯度内作用的主要因子不同,在不同地方有其局部的小环境,如局部变化的光照等[34]。(3)取样误差。实验过程中由于采样时存在误差,同样会模糊性状间关系。
4.2 气孔性状沿海拔梯度的变化
气孔性状对海拔梯度变化的响应,实际是对海拔高度引起的环境因子的响应。不同海拔高度处的温度、降水、空气湿度以及土壤含水量等都存在差异,因此气孔性状也表现出随海拔变化而变化的规律[35]。
关于气孔性状与环境因子之间的关系,前人进行了大量研究,但结果不尽一致[17,36],这可能是由于不同植物的自身特征以及实验过程中的环境不同所致,也可能是未充分考虑到各个环境因子的相互作用对气孔的影响。本研究中土壤湿度与气孔长度之间存在显著负相关关系(P<0.05),但在干旱胁迫条件下,气孔长度减少更具有适应意义。Hetherington 和Woodward[6]提出小气孔具有较短的反应时间,能够快速打开和关闭来应对环境变化,同时,小气孔往往与高气孔密度相连,这能够快速增加气孔导度,使得叶片在有利条件下快速获得较高的气孔导度,当环境不利时能够快速降低气孔导度,叶片能够更好地储存水分,在环境胁迫时期能够保持更加稳定的水分供应。其他土壤因子(土壤总碳、总磷等)与气孔性状的关系在本研究中不显著,可能是研究区内土壤理化性质变化不明显,对气孔性状作用不显著。
降水量虽然对SL与SW变化具有显著影响,但解释量较少,可能是因为影响SL和SW变化的环境因子,除了本文所涉及的以外,还受其他环境因子的影响[10,17,37],有待进一步的探讨和研究。本研究中,空气相对湿度是影响栎属树种气孔导度的主导因子。可以推断,当空气湿度较低或出现干旱胁迫时,叶片从空气中吸收的水分减少,为保持体内水分平衡,则相应地降低气孔导度,减少水分向体外扩散,即气孔导度受植物吸水速率与蒸腾速率平衡的反馈制约[38],是对干旱胁迫环境的一种适应策略。
表现型的可塑性是植物为适应环境变化在生理和形态上发生的变化,并被认为具有生态适应对策的意义[39]。高的可塑性常常与对环境具有高的潜在适应能力相关[40]。本研究中,相比栓皮栎、槲栎和辽东栎,锐齿栎气孔性状的可塑性最大,表明锐齿栎具有更宽的环境适应能力。此外,锐齿栎气孔性状沿海拔梯度表现为非线性特征,可能是因为随着海拔的升高,温度、湿度降低,使环境中的水分不易被树种所吸收导致水分胁迫,适度干旱将增加树种的气孔密度并减小气孔大小,这种小而多的气孔可以适应多变化环境,对水分调节非常有效[34]。但随着干旱的进一步加强,超出锐齿栎适应干旱胁迫影响的范围,气孔密度将显著下降[26],气孔导度也随之降低。因此在约1600 m海拔处是锐齿栎生长的最优地带,具有更高的生长潜力[13,41]。
5 结论
太白山4种栎属树种从整体而言气孔性状间变化特征有一定规律,但不同树种对生境的适应特征存在差异。其次,在4种栎属树种中,锐齿栎对于环境变化具有更强的适应能力。因而在太白山北坡,锐齿栎的分布范围最广。此外,空气相对湿度是影响栎属树种气孔导度的主导因子,但在气孔对环境因子响应高度复杂性的野外自然条件下,各种环境因子对气孔导度存在综合作用,因此,该结果仅能粗略地反映栎属树种叶片气孔导度与环境因子之间的相互关系。
猜你喜欢
农业工程学报(2022年3期)2022-04-16
干旱地区农业研究(2022年1期)2022-01-28
生态学报(2021年6期)2021-04-25
农业工程学报(2021年3期)2021-04-15
节水灌溉(2020年9期)2020-09-16
南方农业·下旬(2020年1期)2020-06-01
装备制造技术(2019年12期)2019-12-25
科技创新与品牌(2019年12期)2019-02-06
现代园艺(2019年24期)2019-01-07
现代农业科技(2009年21期)2018-09-03
