
漓江河岸带枫杨群落主要木本植物种间联结与相关分析
2018-11-14 12:46刘润红李娇凤荣春艳梁士楚杨瑞岸刘星童曾惠帆苏秀丽袁海莹傅桂焕吴燕慧
生态学报 2018年19期
刘润红,姜 勇,常 斌,李娇凤,荣春艳,梁士楚,杨瑞岸,刘星童,曾惠帆,苏秀丽,袁海莹,傅桂焕,吴燕慧
广西师范大学珍稀濒危动植物生态与环境保护教育部重点实验室/广西师范大学生命科学学院,桂林 541006
植物群落是由共存物种在一定生境条件下相互依存、相互竞争和协同进化并与生境相互作用而形成的有机结合体[1- 2]。种间关系是指群落内各种群之间的相互关系,是植物种群间相互联系和相互影响的综合反映,决定了群落的组成结构、功能和动态特征[3]。种间关系的分析方法主要包括种间联结性和种间相关性两个方面。种间联结(Interspecific association)是指不同物种在空间分布上的相互关联性,反映了物种在不同生境中相互影响、相互作用所形成的有机联系,表征了物种间相互吸引或排斥的属性[4]。种间联结通常依据物种存在或不存在二元数据,定性地检验物种之间是否存在关联;而种间相关则是以物种的数量数据(如多度、盖度和重要值等)为依据,体现的是一种定量的关系[5-6]。因此,研究种间联结和相关性,有助于揭示群落的类型、结构、功能和演替趋势,以及植物与环境的关系,从而为植被的恢复与重建、森林的经营管理和生物多样性保护等提供科学理论依据[3]。
枫杨(pterocaryastenoptera)为胡桃科(Juglandaceae)枫杨属(Pterocarya)落叶乔木,广泛分布于我国亚热带和暖温带地区。枫杨喜光,喜潮湿,抗干旱,根系发达,抗风耐火,较耐水淹和土壤贫瘠,生长迅速,适应性强,存活寿命较长,具有防洪护堤、净化水质、绿化环境等重要生态功能,已被广泛用于护堤护岸和造林绿化中[7]。目前,国内外关于枫杨的研究已见许多报道,主要集中在枫杨的群落学研究[8]、灭螺等功能作用[9]、生物量[10]、生物学特性[11]、逆境条件下的生理生态特征[12]、繁殖与栽培[13]、病虫害防治技术[14]、散生木优树选择[15]、育苗技术[16]等方面,但迄今尚未见关于枫杨群落种间关系的研究报道。
河岸带是指位于高低水位之间的河床以及高水位之上直至河水影响完全消失的地带[17]。漓江河岸带是漓江水生生态系统和陆地生态系统之间进行物质传输、能量转化、信息交换和生物流的重要廊道、过滤器和屏障,具有独特的生态系统结构和服务功能,对维持漓江生态系统健康起着极为重要的作用,为漓江流域社会经济和生态的可持续发展提供了有力支撑。枫杨群落作为漓江河岸带植被主要群落类型之一,在涵养水源、净化水质、调节气候、防洪固堤、保护生物多样性和维持河岸带生态平衡等方面均具有重要的作用。然而,近年来,随着旅游业和工农业的快速发展,漓江河岸带枫杨林受人为干扰和破坏日益严重,枫杨林被大量砍伐,枫杨在许多河段已不复存在,导致大量种质资源逐渐丧失,开展枫杨群落的保护研究显得十分紧迫。然而,目前漓江河岸带枫杨群落的研究仍处于探索阶段,其种间联结及相关性研究尚属空白,不利于全面认识其群落结构特征。因此,开展漓江河岸带枫杨群落的种间联结及相关性研究具有十分重要的意义。鉴于此,本研究在野外群落学调查的基础上,基于2×2联列表,采用方差比率(VR)、χ2检验、Pearson相关系数和Spearman秩相关系数检验方法,对漓江河岸带枫杨群落中乔木层和灌木层主要木本植物的种间联结及相关性进行分析,以期揭示该区域枫杨群落的种间关系、群落结构与动态,从而为漓江河岸带植被恢复与重建、森林经营管理、生物多样性保护等提供科学理论依据。
1 材料与方法
1.1 研究区概况
研究区位于桂林市兴安县溶江镇至桂林市区的漓江河岸带上游河段,全长43 km,地理坐标为110°18′—110°27′E,25°18′—25°32′N。该区域属于中亚热带湿润季风气候区,四季分明,气候温和,无春旱,雨热基本同期,全年光照充足,年平均日照时数为1670 h,年平均气温17.8—19.1℃,最冷1月平均气温8℃,最热8月平均气温28℃,年降雨量1814—1941 mm,年蒸发量1377—1857 mm,年平均相对湿度为73%—79%,全年无霜期为309 d[18]。漓江属于典型的雨源型山区河流,径流为降雨所形成,年径流量比较丰富,但年内分配极不均匀,汛期为3—8月,径流量占年径流总量的80%,其中5—6月占年径流量的40%,9—次年2月为枯水期,最枯月1月径流量仅占年径流量的2%[19]。土壤以山地黄壤为主,土层浅薄,厚度分布不均,质地粗,石砾含量高,呈弱酸性[20]。该区域枫杨群落层次结构清晰,乔木层以枫杨为建群种,伴生有朴树(Celtissinensis)、阴香(Cinnamomumburmannii)、乌桕(Sapiumsebiferum)、桑(Morusalba)等物种;灌木层主要有石榕树(Ficusabelii)、空心泡(Rubusrosaefolius)、八角枫(Alangiumchinense)、白饭树(Flueggeavirosa)、萝芙木(Rauvolfiaverticillata)、细叶水团花(Adinarubella)、牡荆(Vitexnegundovar.cannabifolia)等,同时存在一定数量的枫杨、朴树、阴香、桑等乔木幼苗;草本层物种丰富,主要有海岛苎麻(Boehmeriaformosana)、马兰(Kalimerisindica)、竹叶草(Oplismenuscompositus)、柔枝莠竹(Microstegiumvimineum)、麦冬(Ophiopogonjaponicus)、红花酢浆草(Oxaliscorymbosa)、酢浆草(Oxaliscorniculata)、天胡荽(Hydrocotylesibthorpioides)、土牛膝(Achgranthesaspera)等;此外,层间及林下层存在少量的鸡矢藤(Paederiascandens)、乌蔹莓(Cayratiajaponica)、葎草(Humulusscandens)、络石(Trachelospermumjasminoides)、薜荔(Ficuspumila)、葛藟葡萄(Vitisflexuosa)等。
1.2 群落学调查
在充分现场全线踏查的基础上,本研究根据枫杨群落的类型、结构以及生境等特点,选取漓江河岸带上游受人为干扰较轻且发育完好的枫杨群落作为研究对象,建立了20个20 m×20 m的典型样方,其中灵川县灵川镇阳家村和南昌村分别为6、4个、桂林市叠彩区大河乡南洲村和大河村分别为4、6个,各样方基本情况见表1。乔木样方大小为20 m×20 m,在此基础上将每个样方划分成4个10 m×10 m的小样方进行灌木调查。对样方内所有胸径(DBH)≥ 2.5 cm的乔木植株进行每木调查,测量并记录种名、相对坐标、高度、胸径、冠幅等;对灌木层内所有灌木个体(包括胸径≤2.5 cm的乔木幼树)记录种名、相对坐标、基径、高度和冠幅等。同时,记录每个样方的土壤类型、海拔、经纬度、距河距离、砾石裸露率、砾石直径、干扰程度等生境特征数据。
1.3 数据分析与统计
1.3.1 总体联结性检验
本文分别选取漓江河岸带枫杨群落乔木层和灌木层中重要值较高的8种和18种主要物种(表2)进行种间联结与相关性分析。群落的总体联结性采用Schluter[21]提出的根据物种在样方中出现-不出现二元数据的零关联模型计算出的方差比率(VR)来测定,并通过统计量W检验总体关联性是否显著。其具体分析方法如下,首先假设群落内各物种间无显著关联,根据下列公式逐步计算VR值:
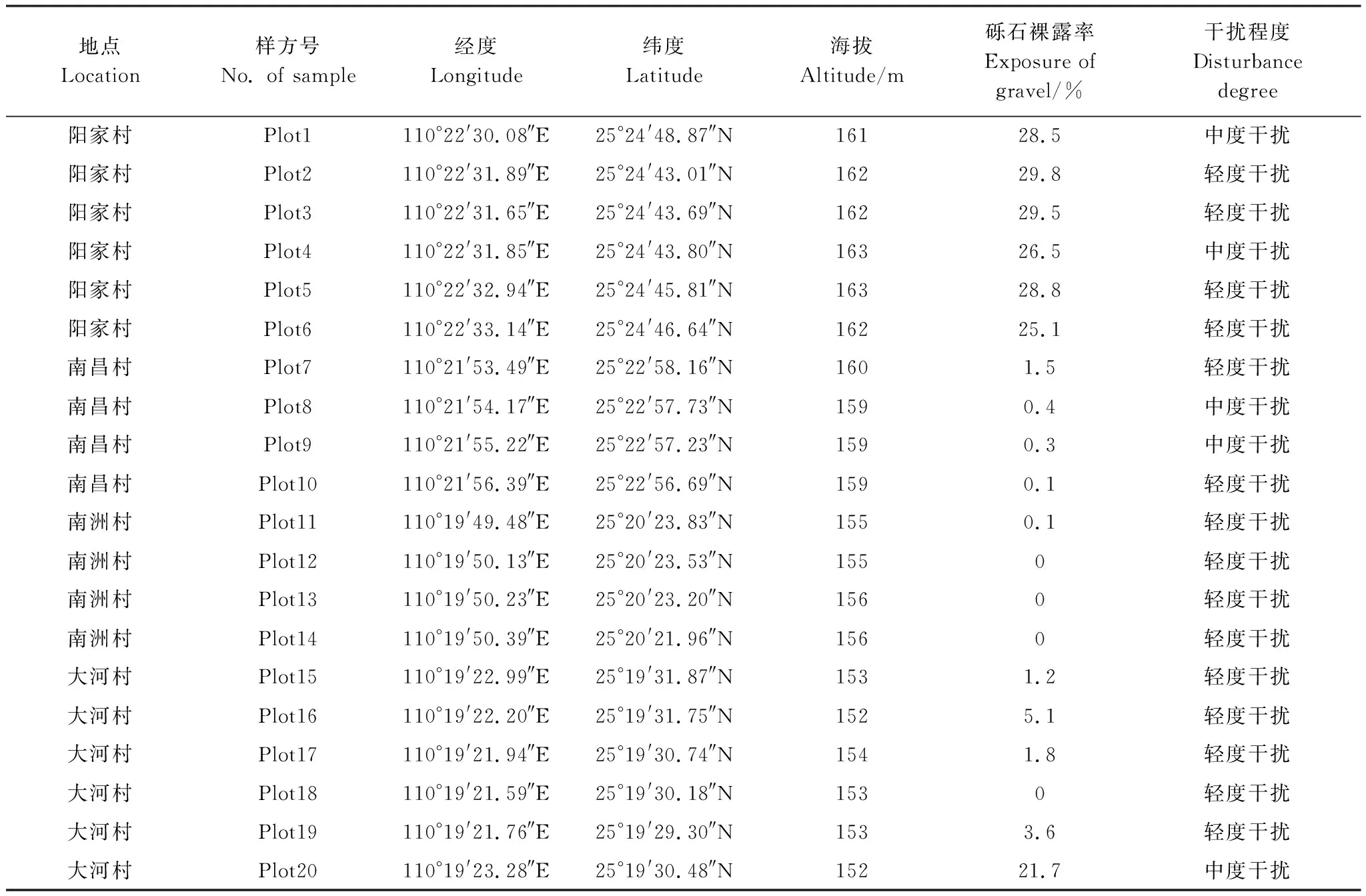
表1 漓江河岸带枫杨群落20个样方的基本情况
Pi=ni/N


1.3.2 种间联结性分析
采用χ2统计量对种间联结性进行定性研究,以确定实测值与在机率基础上预期值之间偏差的显著程度。基于2×2列联表的χ2统计量来判断种对间的联结性,在进行χ2检验前,首先需根据物种在20个样方中是否存在,将原始数据转换成0,1形式的二元数据矩阵,0表示物种在样方中不存在,1表示物种在样方中存在。根据上述原始数据矩阵,将主要种群所有种对的定性数据列入2×2联列表,计算a、b、c、d的值。由于本研究的取样方法为非连续性取样,所以非连续性数据χ2需用Yates的连续校正公式测度[22]。公式如下:

表2 乔木层和灌木层主要物种种名及缩写
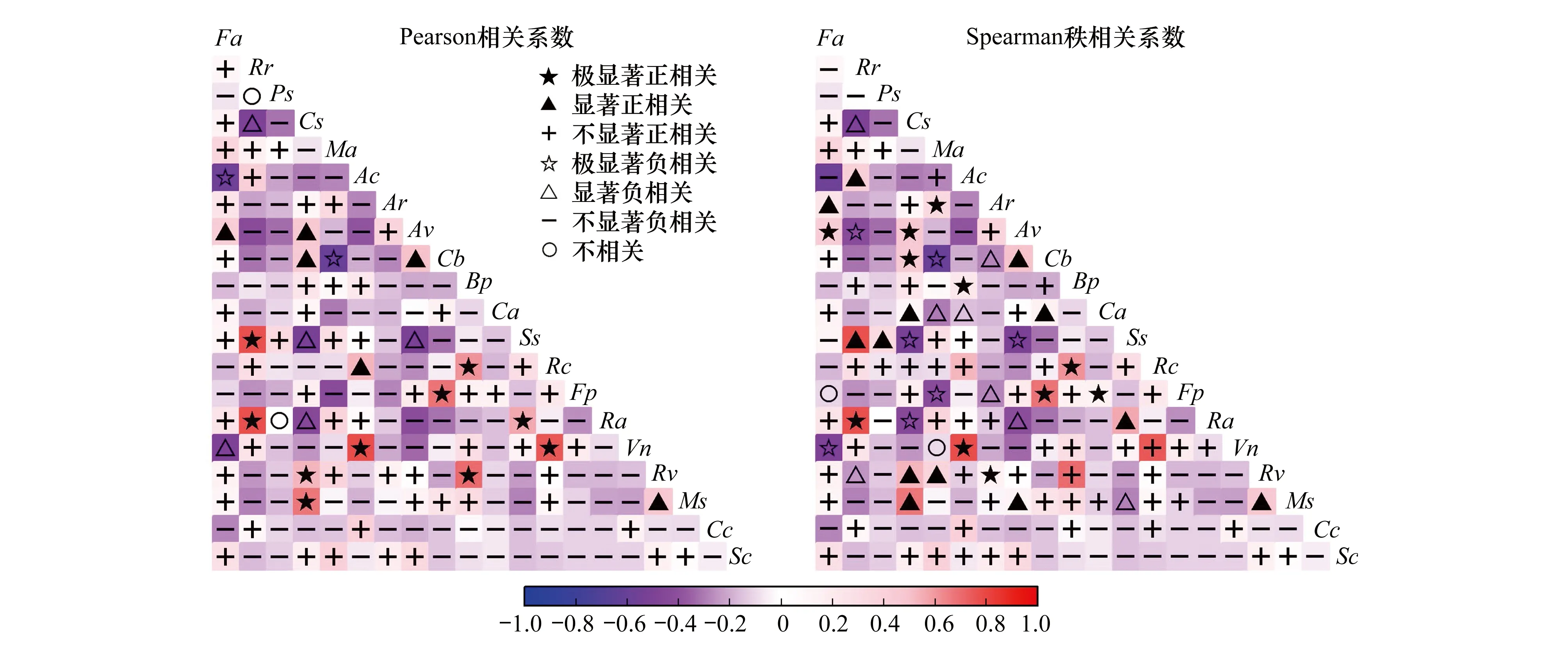
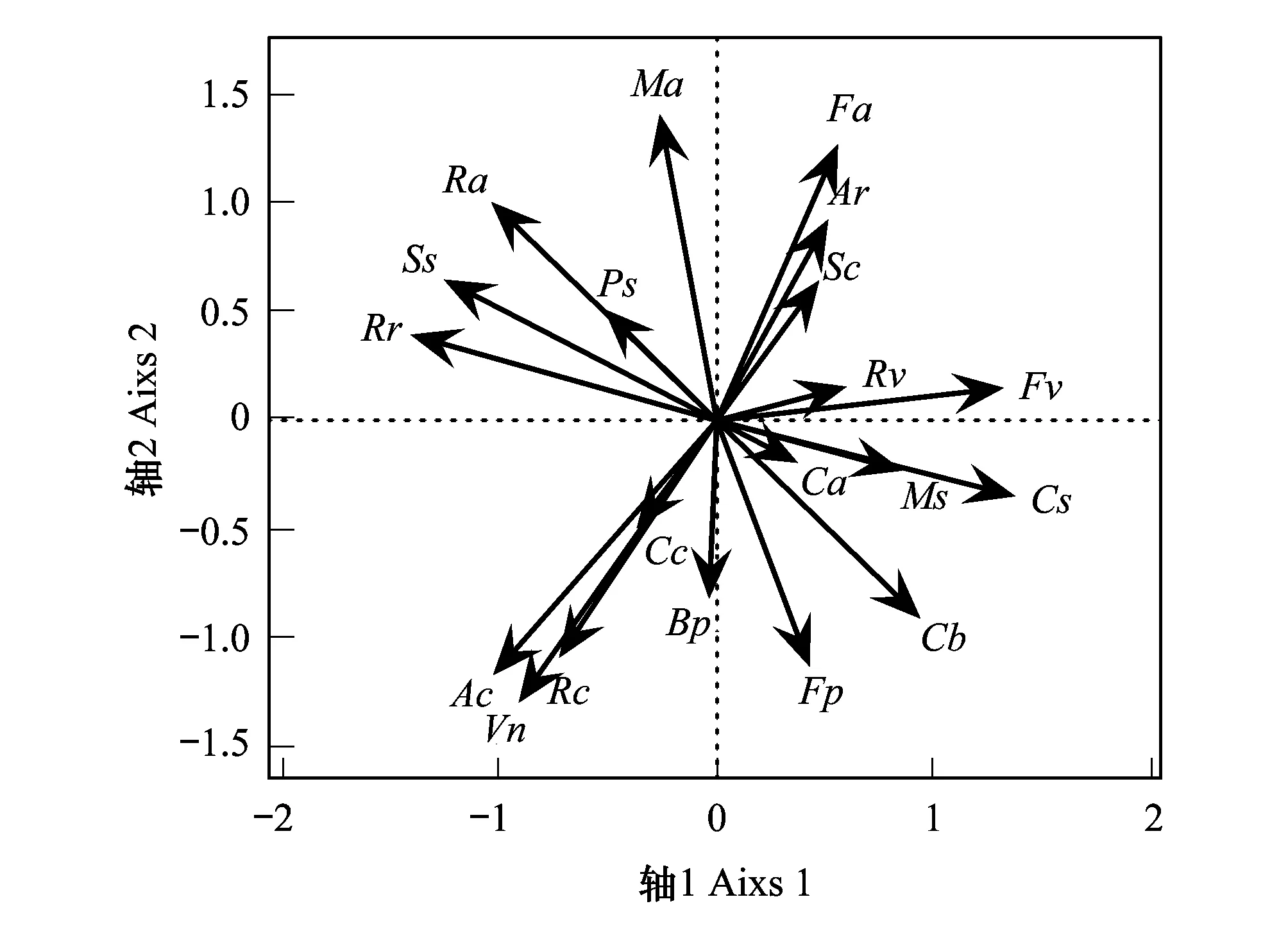
式中,N为总样方数,a为2个物种均存在的样方数,b、c分别为2个物种单独存在的样方数,d为2个物种均不存在的样方数。种间关联性通常可分为正关联和负关联2类,设V=((a+d)-(b+c))/(a+b+c+d),若ad>bc,则V>0,种间关联性为正联结;若ad 当χ2<3.841(P>0.05)时,种对间相互独立,即种对间联结不显著; 当3.841<χ2<6.635(0.01≤P≤0.05)时,种对间联结性显著; 当χ2>6.635(P<0.01),种对间联结性极显著[23]。 1.3.3 种间相关性分析 种间联结性的χ2检验仅定性描述了物种间的关联性是否显著,并不表示那些不显著的种对间就不存在关联性,且χ2检验无法判断联结强度的大小,无法清楚地反映种间关联性的差异性[24]。与之相比,Pearson相关系数检验和Spearman秩相关系数检验则是基于数量数据,能够清楚地反映物种间的线性关系,这2个相关系数检验可以对χ2检验进行有效地补充和完善。因此,在进行种间关联性χ2检验后,需进一步利用Pearson相关系数检验和Spearman秩相关系数检验测定其关联强度。本研究利用乔木层、灌木层主要种群的多度作为Pearson相关系数和Spearman秩相关系数的数量指标,对种对间的线性关系进行定量分析它们同时出现的可能性。 Pearson相关系数计算公式如下: Spearman秩相关系数是首先需要将多度向量转化成秩化向量,然后将秩化向量代入公式计算得出。Spearman秩相关系数计算公式如下: 式中,rp(i,j)分别是种i与种k之间在j样方中的Spearman秩相关系数,N为总样方数,xij和xkj分别为种i和种k在j样方中的多度值的秩。 重要值计算在Excel 2016中完成,种间联结和相关性分析在R 3.4.3软件中实现,其中χ2检验、Pearson相关系数和Spearman秩相关系数检验结果分别由spaa包中的sp.assoc()、sp.pair()函数计算得出,Pearson相关系数和Spearman秩相关系数显著性通过psych包中的corr.test()函数进行检验,PCA分析在vegan包rda()函数中进行。 表3枫杨群落各层次主要种群种间的总体关联性 Table3OverallinterspecificassociationsamongmainplantpopulationsofdifferenthierarchicallevelsofPterocaryastenopteracommunityinriparianzoneofLijiangRiver 层次 Levels方差比率Variance ratio(VR)检验统计量WTest statistics Wχ2临界值χ2 threshold (x20.95,x20.05)检验结果Results乔木层 Tree layer1.1222.42(10.85,31.41)不显著正关联灌木层 Shrub layer1.0621.21(10.85,31.41)不显著正关联乔木和灌木层Combined tree and shrub lay-er1.4528.99(10.85,31.41)不显著正关联 2.2.1 乔木层种群种间关联性 χ2检验统计显示(图1和表4),乔木层28个种对中,无极显著关联种对,朴树与阴香、樟与腺柳2对为显著正关联种对,占总种对数的7.14%;枫杨与樟、枫杨与腺柳2对为显著负关联种对,占总种对数的7.14%;无显著关联或无关联种对有24对,占85.71%,种对间关联较弱,各物种间呈独立分布格局。 表4乔木层种群的χ2检验、Pearson相关检验和Spearman秩相关检验比较 Table4Thecomparisonofχ2-testcoefficient,Pearson′scorrelationcoefficientandSpearman′srankcorrelationcoefficientoftreelayerplantspecies 检验方法Test methods正联结(相关) Positive association(correlation)负联结(相关) Negative association(correlation)极显著Distinctlysignificant(P<0.01)显著Significant(P≤0.05)不显著Not significant(P>0.05)总数Sum极显著Distinctlysignificant(P<0.01)显著Significant(P≤0.05)不显著Not significant(P>0.05)总数Sum无联结(相关) No association (correlation)χ2检验χ2 -test0(0)2(7.14)13(46.43)15(53.37)0(0)2(7.14)7(25)9(32.14)4(14.28)Pearson's相关系数Pearson's correlation coefficient3(10.71)2(7.14)5(17.86)10(35.71)0(0)0(0)18(64.29)18(64.29)0(0)Spearman秩相关系数Spearman's rank correlation coefficient2(7.14)3(10.72)6(21.43)11(39.29)0(0)1(3.57)16(53.57)17(60.71)0(0) 2.2.2 灌木层种群种间关联性 χ2检验显示(图2和表5),153对灌木层主要种对中,阴香与琴叶榕呈极显著正关联,占总种对数的0.65%;朴树与白饭树、朴树与阴香、白饭树与风车子、白饭树与柳叶润楠、风车子与琴叶榕5对为显著正关联,占总种对数的3.27%;枫杨幼树与柳叶润楠、朴树幼树与粗叶悬钩子、白饭树与乌桕幼树、白饭树与粗叶悬钩子、乌桕幼树与柳叶润楠5对为显著负关联,占总种对数的3.27%;无显著关联或无关联种对有142对,占92.81%,种对间关联较弱,各物种间呈独立分布格局。 图1 乔木层种群间种间关联性的χ2检验半矩阵图 Fig.1 Semi-matrix of interspecific correction χ2-test of association of main population in tree layer Ps:枫杨,Pterocarya stenoptera;Cs:朴树,Celtis sinensis;Cb:阴香,Cinnamomum burmanni;Ss:乌桕,Sapium sebiferum;Ma:桑,Morus alba;Bp:构树,Broussonetia papyrifera;Cc:樟,Cinnamomum camphora;Sc:腺柳,Salix chaenomeloides 图2 灌木层18个主要种群间χ2检验半矩阵图 Fig.2 Semi-matrix of interspecific association χ2 correction test of 18 main species in shrub layerFa:石榕树,Ficus abelii;Rr:空心泡,Rubus rosaefolius;Ac:八角枫Alangium chinense;Ar:细叶水团花,Adina rubella;Fv:白饭树,Flueggea virosa;Ca:风车子,Combretum alfredii;Rc:小果蔷薇,Rosa cymosa;Fp:琴叶榕,Ficus pandurata;Ra:粗叶悬钩子,Rubus alceaefolius;Vn:牡荆, Vitex negundo var. cannabifolia;Rv:萝芙木,Rauvolfia verticillata;Ms:柳叶润楠,Machilus salicina Table5Thecomparisonofχ2-testcoefficient,Pearson′scorrelationcoefficientandSpearman′srankcorrelationcoefficientofshrublayermainplantspecies 检验方法Test methods正联结(相关) Positive association(correlation)负联结(相关) Negative association(correlation)极显著Distinctlysignificant(P<0.01)显著Significant(P≤0.05)不显著Not significant(P>0.05)总数Sum极显著Distinctlysignificant(P<0.01)显著Significant(P≤0.05)不显著Not significant(P>0.05)总数Sum无联结(相关) No association (correlation)χ2检验χ2-test1(0.65)5(3.27)58(37.91)64(41.83)0(0)5(3.27)63(41.17)68(44.44)21(13.73)Pearson相关系数Pearson's correlation coefficient8(5.23)6(3.92)49(32.03)63(41.18)1(0.65)6(3.92)83(54.25)90(58.82)0(0)Spearman秩相关系数Spearman's rank correlation coefficient11(7.19)12(7.84)51(33.33)74(48.36)7(4.58)12(7.84)58(37.91)77(50.33)2(1.31) 2.2.3 乔木和灌木层种群种间关联性 χ2检验统计显示(图3和表6),乔木和灌木层190对种对中,无极显著关联种对;有11对呈显著正关联,占总种对数的5.78%,分别为石榕树与枫杨、石榕树与樟、石榕树与腺柳、枫杨与樟、枫杨与腺柳、朴树与白饭树、朴树与阴香、白饭树与风车子、白饭树与柳叶润楠、阴香与琴叶榕、风车子与琴叶榕;有4对为显著负关联,占总种对数的2.11%,分别为朴树与粗叶悬钩子、白饭树与乌桕、白饭树与粗叶悬钩子、樟与腺柳;无显著关联或无关联种对有175对,占92.11%,种对间关联较弱,各物种间呈独立分布格局。 表6乔木和灌木层种群的χ2检验、Pearson相关检验和Spearman秩相关检验比较 Table6Thecomparisonofχ2-testcoefficient,Pearson′scorrelationcoefficientandSpearman′srankcorrelationcoefficientofcombinedtreeandshrublayerplantspecies 检验方法Test methods正联结(相关) Positive association(correlation)负联结(相关) Negative association(correlation)极显著Distinctlysignificant(P<0.01)显著Significant(P≤0.05)不显著Not significant(P>0.05)总数Sum极显著Distinctlysignificant(P<0.01)显著Significant(P≤0.05)不显著Not significant(P>0.05)总数Sum无联结(相关) No association (correlation)χ2检验χ2-test0(0)11(5.78)80(42.11)91(47.90)0(0)4(2.11)84(44.21)88(44.32)11(5.79)Pearson相关系数Pearson's correlation coefficient10(5.26)6(3.16)55(28.95)71(37.37)2(1.05)5(2.63)110(57.90)117(61.58)2(1.05)Spearman秩相关系数Spearman's rank correlation coefficient11(5.79)13(6.84)62(32.63)86(45.26)7(3.69)8(4.21)87(45.79)102(53.69)2(1.05) 图3 乔木和灌木层20个主要种群间χ2检验半矩阵图 Fig.3 Semi-matrix of interspecific association χ2 correction test of 20 main species in combined tree and shrub layer 2.3.1 乔木层种群种间相关性 乔木层种群Pearson相关检验如图4和表4所示,在所有28个种对中,有3对为极显著正相关,2对呈显著正相关,总共有5对,占总对数的17.85%;无呈显著负相关和极显著负相关的种对出现,检验显著率为17.85%。其中,种间相关性不显著的种对占绝大部分,种间相关性不强,这同χ2检验的结果相同。 乔木层种群Spearman秩相关检验如图4和表4所示,在所有28个种对中,有2对为极显著正相关,3对为显著正相关,总共有5对,占总对数的17.85%;无极显著负相关种对,显著负相关种对1对,检验显著率为21.43%。其中,种间相关性不显著的种对占绝大部分,种间相关性不强,这同χ2检验和Pearson相关检验的结果相似。 图4 乔木层种群间的Pearson相关系数和Spearman秩相关系数半矩阵图Fig.4 Semi-matrix of Pearson′s correlation coefficients and Spearman′s rank correlation coefficients of main population in tree layer 2.3.2 灌木层种群种间相关性 灌木层主要种群Pearson相关系数检验结果如图5和表5所示,在所有153个种对中,呈极显著正相关的种对有8对,呈显著正相关的种对有6对,总共有14对,占总种对数的9.15%;有1对呈极显著负相关,有6对呈显著负相关,总共有7对,占总种对数的4.57%,检验显著率为13.72%。其中,大部分种对的种间相关不显著,种间相关性不强,这与乔木层和灌木层χ2检验的结果一致。 灌木层主要种群Spearman秩相关系数检验结果如图5和表5所示,在所有153个种对中,有11对呈极显著正相关,有12对呈显著正相关,总共有23对,占总对数的15.03%;呈极显著负相关的种对有7对,呈显著负相关的种对有12对,总共有19对,占总对数的12.42%,检验显著率为27.45%。其中,大部分种对的种间相关不显著,种间相关性不强,这同乔木层以及灌木层χ2检验和Pearson相关系数检验的结果相一致。 图5 灌木层种群间的Pearson相关系数和Spearman秩相关系数半矩阵图 Fig.5 Semi-matrix of Pearson′s correlation coefficients and Spearman′s rank correlation coefficients of main population in shrub layer 2.3.3 乔木和灌木层种群种间相关性 乔木和灌木层主要种群Pearson相关系数检验结果如图6和表6所示,在所有190对种对中,呈极显著正相关的种对有10对,呈显著正相关的种对有6对,总共有16对,占总种对数的8.42%;有2对呈极显著负相关,有5对呈显著负相关,总共有7对,占总种对数的3.68%,检验显著率为12.11%。其中,大部分种对的种间相关性不显著,相关性不强,这与乔木和灌木层χ2检验的结果一致。 乔木和灌木层主要种群Spearman秩相关系数检验如图6和表6所示,在所有190个种对中有11对呈极显著正相关,有13对呈显著正相关,总共有24对,占总对数的12.63%;呈极显著负相关的种对有7对,呈显著负相关的种对有8对,总共有15对,占总对数的7.89%,检验显著率为20.53%。其中,大部分种对的种间相关不显著,种间相关性不强,这同乔木和灌木层χ2检验和Pearson相关系数检验的结果相一致。 图6 乔木和灌木层种群间的Pearson相关系数和Spearman秩相关系数半矩阵图 Fig.6 Semi-matrix of Pearson′s correlation coefficients and Spearman′s rank correlation coefficients of main population in combined tree and shrub layer 生态种组是群落中生态习性相似的物种的组合,能够更准确地表达种群和群落的相互关系和种群对环境资源和主导生态因子的适应策略。本研究综合2 × 2联列表、χ2检验、Pearson相关系数、Spearman秩相关系数检验的结果,结合各物种对环境因子的不同生态适应特征,运用PCA排序分析,将乔木和灌木主要物种大致分为4大生态种组,结果如图7所示。 图7 乔木和灌木层主要物种PCA二维排序图 Fig.7 Ordination of main plant species in combined tree and shrub layer on the first and second principal component axis 第一生态种组:石榕树(Fa)、细叶水团花(Ar)、腺柳(Sc)、萝芙木(Rv)、白饭树(Fv),它们都位于第一主成分轴右侧第二主成分轴上方,耐阴喜湿,多生长于靠近漓江河岸边的林下,种间关联基本呈不显著或显著的正联结,生态习性与生境要求基本一致。第二生态种组:空心泡(Rr)、乌桕(Ss)、粗叶悬钩子(Ra)、枫杨(Ps)、桑(Ma),它们都分布在第一主成分轴的左侧第二主成分轴上方,该生态种组的植物均具有喜阳等特点。第三生态种组:八角枫(Ac)、牡荆(Vn)、小果蔷薇(Rc)、樟(Cc)、构树(Bp),分布在第一主成分轴的左侧第二主成分轴下方,该生态种组的植物均具有喜阳又略耐荫等特点,两两之间多呈不显著或显著正联结。第四生态种组:琴叶榕(Fp)、阴香(Cb)、风车子(Ca)、柳叶润楠(Ms)、朴树(Cs),分布在第一主成分轴的右侧第二主成分轴下方,该生态种组的植物均具有稍耐荫等特点,种间关联基本呈不显著或显著的正联结,生态习性和生境要求基本一致。 种间联结性与群落演替相互关联,种群间总体关联性体现了群落演替的阶段及其稳定性,群落稳定性是种间联结性的表达形式[25]。种间联结性是对一定时期内群落组成物种间相互关系的静态描述,这种关系不仅包括空间分布关系,同时也隐含着物种之间的功能,它随群落的发展而变化,在一定程度上反映物种间的相互关系及其与环境间的关系[26- 27]。然而,当群落逐渐向顶极群落发展时,其种间联结性如何发生变化,学者说法不一。如,张明霞等[28]认为随着演替进展,群落结构及物种组成将逐渐趋于完善和稳定,种对间正联结比例增加,以求得物种间的稳定共存;而黄世能等[29]认为随着演替进展,群落中的物种总体趋于无关联。本研究发现漓江河岸带枫杨群落乔木层、灌木层以及乔木和灌木层总体种间关系均呈不显著正关联,呈不显著关联的种对占绝大部分,种对间的关联性较弱,表明漓江河岸带枫杨群落正处于趋于相对稳定的顶级群落演替阶段,群落中的物种组成和群落结构也趋于稳定和完善,各种群之间都能占据有利于自己的生态位,相互依赖和相互竞争大为减弱,物种间的联结较为松散,独立性相对较强。这可能与漓江河岸带枫杨群落的环境条件、目前的发展阶段、物种自身的生物、生态学特性以及外界的干扰有关。漓江河岸带受江水的持续冲刷和淤积,地形地貌复杂多变,地表支离破碎,微生境类型复杂多样,组合、分布随机,不同尺度生境异质性的相互耦合加剧了漓江河岸带生境的分异和复杂程度,由于植物对生境的生态适应性存在差异,造成物种在地理空间分布上的差异,种间联系多被微生境所割裂,各种群相遇的概率变低,造成种对间独立性相对较强。同时,群落的发生、发展和演替过程既是环境对物种筛选的过程,也是物种对环境的适应过程,在生境过滤和种间竞争的共同作用下,群落中各物种相互适应和协同进化,各种群在物种的形态、结构和生理等方面出现了许多趋向于相互弥补、相互配合的特征,如高度的差异等,导致种间生态位发生一定程度的分化,各个物种都占据着有利的位置,和谐共处,相互依赖和相互竞争大为降低和减弱,所以大多数种对联结程度不强,关系松散,独立性强[28]。因此,种间松散的联结关系可能是漓江河岸带枫杨群落物种共存机制之一,而不同尺度上生境异质性及其耦合效应和植物的生理生态学适应策略是阐述漓江河岸带枫杨群落物种共存机制的基础。此外,外界干扰也会降低群落结构的稳定性,漓江河岸带枫杨群落受自然和人为干扰较严重,给植物的生长带来一定负面影响。 种间联结性是各个物种在不同生境条件下相互影响、相互作用而形成的有机联系,它反映了物种间的相互关系及其对环境因子的生态适应性[30]。种间联结理论认为,正联结是由于物种之间互相依赖或者是由于在异质环境中,物种之间对环境条件具有相同或相似的适应策略,导致各物种之间相互兼容,互相促进生长,常常相伴出现[31]。在相同环境条件下,植物种对的正联结可能在一定程度上表明相互作用的存在对一方或双方都是有利的,如互惠共生和资源划分的互补等,体现了植物利用资源方式的相似性和生态位的重叠性[27]。如,枫杨与乌桕均为乔木层喜阳物种,分布在乔木层的第一层,空心泡和粗叶悬钩子均为蔷薇科喜阳、耐土壤贫瘠灌木物种,由于对光照等环境资源需求较相似,种群间能够共同分享资源,两者生态位产生重叠而呈现极显著正联结。 种对间的负联结是由于生物学特性的不同以及种群间对环境需求的差异性或者在竞争资源时发生相互排斥,可能指示不利于一方或双方的相互作用,如种间竞争、干扰等,体现了物种间的排斥性和生态位分离,能够反映各种群对环境异质性的适应性[32- 33]。如,种对乌桕与白饭树,乌桕是组成漓江河岸带植被的主要乔木树种,常分布在地势较高的河漫滩上,对光照要求较强,而白饭树为较耐荫的灌木物种,多生长在靠近河岸的乔木林下,两者由于对生境要求的差异而产生生态位分离,从而呈现极显著负联结;阴香与乌桕,多位于乔木层,两者对光照等环境条件需求较相似,形成了激烈的竞争,从而呈现显著负联结关系,表现出较强的排斥性。 本研究中Pearson相关系数和Spearman秩相关系数的检验结果显示乔木层和灌木层负联结种对数均多于正联结种对数,但群落的总体联结性却呈现不显著正联结,这一研究结果与简敏菲等[24]对九连山常绿阔叶林乔木主要种群的种间关联性分析结果一致。这是由于本研究结果中的正联结相对数值和具有显著或极显著关联的种对数均高于负联结的相对数值和显著或极显著种对数,说明漓江河岸带枫杨群落在长期演替过程中,群落内主要种群的生态位产生了分化,部分物种间存在有利的相互作用,形成了比较稳定的结构,这可能与枫杨群落内部分物种的生态习性密切关联。建群种枫杨与其他物种的联结性不强,甚至基本趋于负相关,这是由于主要种群间进行自我调节、分配和利用生存空间和资源的结果。一般位于乔木层第1亚层的植物个体,由于占据着自已合适的生态位,与其他物种的联结性不强[34]。 χ2检验、Pearson相关系数检验和Spearman秩相关系数检验的检验结果具有一致性,但也存在一定的差异。这主要是由于χ2检验是基于物种在样方中是否存在的0、1二元矩阵数据作为衡量联结性的标准,虽然计算方法简单,且具有明确的判断标准(P<0.01和P<0.05),但是其难免会损失种间相对多度等信息,仅能定性判断种对间是否存在关联,而不能定量地测度其关联程度的大小[33,35]。Spearman秩相关系数检验结果发现的显著与极显著的种对数基本都多于Pearson相关系数检验,主要是由于Pearson相关系数检验属于参数检验方法,要求物种必须服从连续的正态分布,然而自然界大多数物种均呈集群分布,从而使得Pearson相关系数检验的结果存在一定局限性,而Spearman秩相关系数检验是一种非参数检验方法,对物种的分布形式没有任何要求。因此,Spearman秩相关系数检验的灵敏度较Pearson相关系数检验的高,应用起来也较为方便,可以有效地弥补Pearson相关系数检验的不足。这与前人大多数研究结果一致[4, 36- 37]。因此,在进行种间关系研究时,要以χ2检验为基础,定性测定物种间联结性的显著性,辅以Pearson相关系数和Spearman秩相关系数等方法进行检验,共同分析群落中各种对的种间关系,从而相对全面、准确地反映植物群落的种间联结性。 生态种组是群落中生态习性相似的物种的组合,而群落的种间联结性和相关性反映了群落内各物种由于受微生境影响而表现在空间分布上的相互关系,从而种间联结性的结果可以作为生态种组划分的重要依据[26]。本研究利用PCA排序,综合2 × 2联列表、χ2检验、Pearson相关系数、Spearman秩相关系数检验的结果,结合物种对环境因子的不同生态适应特性,将漓江河岸带枫杨群落主要乔木和灌木物种大致划分为4大生态种组。同一生态种组内各物种之间具有较强的正关联或正相关,因为它们的资源利用方式、生物学特点和生态习性比较相似,对生境具有一致的生态适应性和生态位相互分离。相反,不同生态种组间的种对由于它们的资源利用方式、生物学特点和生态习性存在差异,对生境具有不同的生态适应性和生态位重叠,从而造成各生态种组间的种对呈负关联或负相关。但是,仅从上述几个方面进行生态种组划定,由于某些物种出现的频度较小,从而得出的种间联结性和PCA排序结果也不非常可靠,因此,在以后的生态种组划分研究中要进一步结合土壤理化性质等环境因子、植物生活型和生活习性,以准确进行生态种组的划分。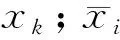
2 结果与分析
2.1 总体关联性


2.2 种间关联性






2.3 相关性分析


2.4 生态种组的划分
3 结论与讨论
3.1 群落演替与种间联结的关系
3.2 种间联结性与相关性
3.3 不同检验方法的对比
3.4 生态种组与种间联结的关系
猜你喜欢
现代农业科技(2022年10期)2022-12-17
河南师范大学学报(自然科学版)(2022年5期)2022-08-08
蔬菜(2018年5期)2018-05-17
乡村科技(2018年8期)2018-02-13
扬子江(2018年1期)2018-01-26
诗潮(2017年2期)2017-03-16
湖南林业科技(2017年1期)2017-02-06
杂草学报(2015年2期)2016-01-04
湖南农业科学(2015年11期)2015-01-10
中国海洋大学学报(自然科学版)(2014年12期)2014-02-28
