
Isolation and callus formation of Gracilariopsis bailiniae(Gracilariales, Rhodophyta) protoplasts*
2018-12-22 07:00CHENHaihong陈海红CHENWeizhou陈伟洲SHIJingyi石经仪CHENZepan陈泽攀ZHANGYi张毅
CHEN Haihong (陈海红), CHEN Weizhou (陈伟洲), SHI Jingyi (石经仪),CHEN Zepan (陈泽攀), ZHANG Yi (张毅)
Marine Biology Institution, Key Laboratory of Marine Biotechnology of Guangdong Province, Shantou University, Shantou 515063, China
Abstract This paper reports the first successful isolation of protoplasts from Gracilariopsis bailiniae and their callus formation. The base solution type, concentration of isolating enzymes, concentration of sorbitol,incubation time, temperature and pH of the enzyme solution were tested to optimize the protoplast yield.The optimized isolation conditions were: 40% base solution 3 (deionized water containing 25 mmol/L MESTris and 25 mmol/L CaCl 2·2H 2 O) and 60% crude Marinomonas sp. YS-70 agarase solution, containing 2%w/v cellulase, 1% w/v macerozyme R-10 and 0.4 mol/L sorbitol, with incubation for 4 h at 28°C and pH 6.5.The highest yield of viable protoplasts, which was obtained in these conditions, was (1.75±0.25)×10 6 cells/g fresh weight. Cell wall regeneration of most protoplasts from G. bailiniae was complete within 60 h and the first division of cells happened after ≥3 days. Two division types were observed in the first division of protoplasts from G. bailiniae— asymmetric division and symmetric division. After the first division, the cells underwent a series of divisions to form callus cell masses.
Keyword: Gracilariopsis bailiniae; enzyme; marine bacterium; protoplast isolation; cell division; callus
1 INTRODUCTION
Gracilariopsisbailiniaeshows fast growth and heat-resistance (Zhong et al., 2014). It is a food for abalones and a raw material for agar extraction that is farmed in Fujian and Guangdong Provinces, China(Hurtado-Ponce, 1992; Pan and Li, 2010).Agarophytes (Gelidium,GracilariaandGracilariopsis) have considerable industrial importance since they are the principal source of raw material for the agar industry worldwide (Zemke-White and Ohno, 1999; Smit, 2004). In recent years,there has been increasing global demand for agar,which is used widely for microbial culture and in the food, health care products, medical and chemical industries, because of its gel, thickening and stabilization properties (Liu et al., 2013). The supply of agarophytes from wild stocks can no longer meet the demand for agar. Thus, large-scale cultivation of agarophytes is required (Gupta et al., 2013). The continuous supply of seed material is a key step for successful aquaculture (Saminathan et al., 2015).However, the limitations of traditional seed production methods for most agarophytes have restricted exploitation of these seaweeds for industrial use(Mantri, 2009). Thus, development of seed stock methods for agarophytes is crucial to achieve largescale production of seedlings.
Protoplasts are living plant cells devoid of cell walls which can be applied in somatic hybridization,proteomics, metabolomics, cybridization and protoclonal variation studies (Fujita and Saito, 1990;Davey et al., 2005; Reddy et al., 2008). Theoretically,a protoplast can lead to the regeneration of one or more plants from a single cell due to the totipotence of plant cells (Huddy et al., 2013). Moreover, many protoplasts can be isolated from a small fragment of algal thallus, so protoplasts can be excellent tools for seed stock production and plant breeding (Gupta et al., 2011; Huddy et al., 2013; Wang et al., 2014). Chen(1998) and Chen and Shih (2000) developed methods for producing stocks of seedlings using protoplasts from green algae (UlvafasciataandMonostroma latissimum). Protoplasts fromMonostromaandPorphyrahave also been successfully tested for seeding and regeneration in laboratory conditions(Kito et al., 1998; Dipakkore et al., 2005; Reddy et al., 2006), and protoplasts from some species ofGracilariaandGracilariopsishave successfully been regenerated to whole thalli (Cheney, 1990; Yan and Wang, 1993; Reddy et al., 2008; Yeong et al., 2008;Wang et al., 2014; Huddy et al., 2015). These studies indicated that using protoplasts as seed stocks for cultivation is feasible. However, methods for protoplast isolation and regeneration of agarophyte species have not been sufficiently established (Wang,1994; Baweja et al., 2009). There are very few reports about protoplast isolation fromGracilariaandGracilariopsisspecies (Reddy et al., 2010; Gupta et al., 2011; Huddy et al., 2013; Wang et al., 2014).
In the present study, we used a marine bacterium,Marinomonassp. YS-70, that can produce agarase.The crude agarase solution from this bacterium mixed with commercial enzymes (cellulase and macerozyme R-10) as a cell wall hydrolase produced good yields of protoplasts fromGracilariopsisbailiniae. This paper reports the optimization of the protoplast isolation conditions and the callus formation of these protoplasts.
2 MATERIAL AND METHOD
2.1 Experimental algae
Thalli ofG.bailiniaewere obtained from the breeding pond of Hainan Ocean and Fisheries Sciences Research Base, Qionghai, Hainan, China. In the laboratory, the seaweed samples were cleaned of mud and epiphytes using a soft brush and filtered seawater. Then, the thalli were maintained in ventilated tanks at 27°C and 40–60 μmol photons/(m2·s) (12 h light:12 h dark) (Zhong et al., 2014).
2.2 Preparation of agarase
Marinomonassp. YS-70, which was isolated from red algae, was used for the preparation of agarase.The bacteria were inoculated into 100 mL of 2216E medium in a 500-mL conical flask. The 2216E medium was composed of 5 g tryptone (OXOID), 1 g yeast extract (OXOID), 0.01 g FePO4(Sangon) and 0.2% agar (OXOID), dissolved in 1 000 mL aged seawater (final pH adjusted to 7.3). After incubating at 26°C for 48 h in a reciprocal shaker (120 r/min), the bacterial solution was centrifuged at 10 444 r/min for 30 min at 4°C. The supernatant was collected and stored at -80°C until use in protoplast isolation.
2.3 Agarase assay
Agarase assay was carried out by estimating the reducing sugar released using the 3,5-dinitrosalisylic acid (DNS) method (Tang, 2012). The activity of agarase was determined at pH 3.5, 4.5, 5.5, 6.5, 7.5,8.5, 9.5 and 10.5. The reaction mixture was prepared from 0.1 mL crude agarase solution and 0.9 mL buffer(0.05 mol/L) containing 20 g/L agar; the buffers used were citric acid (pH 3.5–5.5), phosphate (pH 6.5–7.5),and glycine-NaOH (pH 8.5–10.5). The reaction mixture was incubated at 40°C for 30 min. Then,1 mL of DNS was mixed with 1 mL of the reaction mixture, heated for 5 min in a boiling water bath, and then cooled. The release of reducing sugar was determined by measuring the absorbance at 540 nm against a standard curve for galactose. One unit of agarase activity was defined as the amount of enzyme(mL) that produced reducing sugar equivalent to 1 μg D-galactose per min in these conditions.
2.4 Isolation of protoplasts from G. bailiniae
For the optimization of protoplast isolation, thalli were cleaned with a soft brush and rinsed three times with filtered seawater. After cleaning,G.bailiniaethalli were cut into 2–3 mm long pieces using a sterile scalpel blade in a culture dish and rinsed three times in filtered seawater. Then, approximately 0.5 g of seaweed pieces were incubated in 5 mL of enzyme solution in a 50-mL conical flask placed in the dark on a rotary shaker (90 r/min) for 4 h at pH 6.5. The initial enzyme solution (before various parameters were optimized) contained 1% (w/v) cellulase and 0.8 mol/L sorbitol in 40% base solution (deionized water containing 25 mmol/L 2-(N-morpholino)ethanesulfonic acid-Tris [MES-Tris] and 25 mmol/L CaCl2·2H2O) and 60% crude agarase solution. After incubation, the enzyme mixture was filtered through 45 μm nylon mesh to remove undigested algal pieces,and the remaining filtrate was centrifuged at 1 247 r/min for 8 min at 25°C. After that, 80% of the supernatant was discarded, and the protoplasts were resuspended in 5 mL MES medium containing 0.6 mol/L D-sorbitol and this procedure was repeated twice. Then,protoplast numbers were determined in a blood counting chamber. Each set of conditions was tested with three replicates.
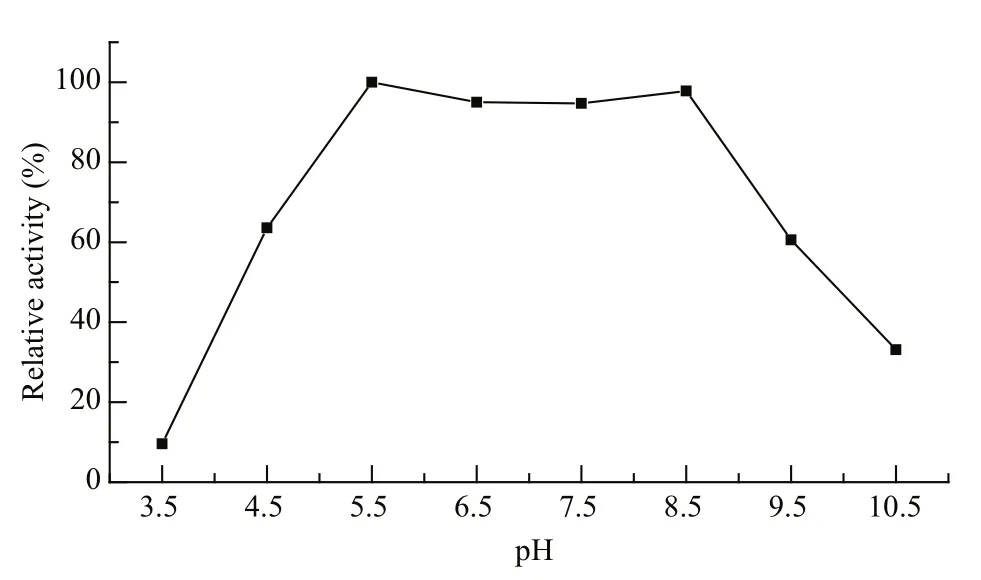
Fig.1 pH-activity curve of Marinomonas sp. YS-70 agarase
For protoplast regeneration experiments, the thalli ofG.bailiniaewere prepared more stringently. First,thallus tips (within 3 cm from the apex) were selected,cleaned with a soft brush and rinsed three times with filtered seawater. After that, the thallus tips were treated with an ultrasonic cleaner (KQ-250DB) at 100% power for 3 min before soaking in sterile seawater containing 1.5% KI for 10 min. Finally, the thallus tips were immersed in sterile seawater containing 0.1 g/L ampicillin sulfate, 0.1 g/L kanamycin sulfate, 0.02 g/L neomycin sulfate and 2 mg/L GeO2for 48 h at 25°C.Then, theG.bailiniaethalli were used to isolate protoplasts, as described above.
2.5 Optimization of protoplast isolation parameters
For protoplast isolation fromG.bailiniae, first, the base solution was optimized from among seawater(base-solution 1), deionized water (base-solution 2),and deionized water containing 25 mmol/L MES-Tris and 25 mmol/L CaCl2·2H2O (base-solution 3). Then,the concentration of cellulase (0%, 1%, 2%, 3% or 4% w/v), macerozyme R-10 (0%, 0.5%, 1% or 1.5%w/v) and sorbitol (0.2, 0.4, 0.6, 0.8 or 1.0 mol/L), and the incubation time (2, 3, 4, 5 or 6 h), temperature (25,28 or 31°C) and enzyme solution pH (5.9, 6.1, 6.3, 6.5 or 6.7), were also optimized, in order. The optimal conditions identified in the proceeding tests were used in subsequent tests.
2.6 Protoplast staining
To confirm the viability of protoplasts, we used 0.5% w/v Evans blue (Biotopped, Beijing, China) to stain protoplasts, which were observed under a light microscope (Zhang et al., 2014). The viable protoplast yields were determined as follows:
Viable protoplast yield=total number of protoplasts×survival rate.

Fig.2 Effect of different base solutions on protoplast yield of G. bailiniae thalli
To confirm true protoplasts lacking a cell wall,0.01% (w/v) Fluorescent Brightener 28 (Sigma) was used to stain protoplasts, and they were observed under a fluorescence microscope (LEICA DMI3000 B) with UV light (Wang et al., 2014).
2.7 Culture of protoplasts
Protoplasts ofG.bailiniaewere dispensed into 2 mL MES medium containing 0.6 mol/L sorbitol in 35 mm×10 mm Petri dishes with a protoplast density of 5×104–1×105cells/mL. Then, protoplasts were cultured at 26°C with a 12 h:12 h light:dark cycle(16 μmol/(m2·s)). After culturing for 2 days, the culture medium was replaced with ½ MES medium containing 0.6 mol/L sorbitol. Then, the ½ culture medium was replaced with MES medium when the protoplasts had been cultured for a total of 4 days.After that, 50% of the culture medium was replaced every 3 days with MES medium.
2.8 Statistical analysis
Analysis of variance was used for the comparison of results in different conditions.P<0.05 was considered significant.
3 RESULT
3.1 Agarase activity assay
Marinomonassp. YS-70 agarase activity was high at pH 5.5 to 8.5 (Fig.1) and the highest enzyme activity was observed at pH 5.5. Lower and higher pH values resulted in lower enzyme activity.
3.2 Effect of the base solution type

Fig.3 Effect of enzyme constituents and concentrations on protoplast yield from G. bailiniae thalli
As Fig.2 shows, the protoplast yield was significantly (P<0.05) increased by using basesolution 3 and the highest protoplast yield((12.5±1.3)×105cells/g fresh weight [f. wt]) was obtained with this treatment. There was no significant difference in the protoplast yield between treatment with base-solution 1 and base-solution 2. The lowest protoplast yield ((1.08±0.59)×105cells/g f. wt) was obtained on treatment with base-solution 1.
3.3 Optimization of cell wall degrading enzyme combination and concentration
As Fig.3a and b show, the protoplast yields fromG.bailiniaethalli were significantly affected by the concentration of added enzymes (P<0.05). The protoplast yield was highest when the enzyme mixture contained 60% crudeMarinomonassp. YS-70 agarase solution, 2% w/v cellulase and 1% w/v macerozyme R-10. A large quantity of protoplasts could be obtained when the mixture of enzymes consisted of agarase and cellulase only (Fig.3a), i.e., when the concentration of macerozyme R-10 was 0%. However, few protoplasts were released fromG.bailiniaethalli when agarase was mixed with macerozyme R-10 without cellulase (Fig.3b).
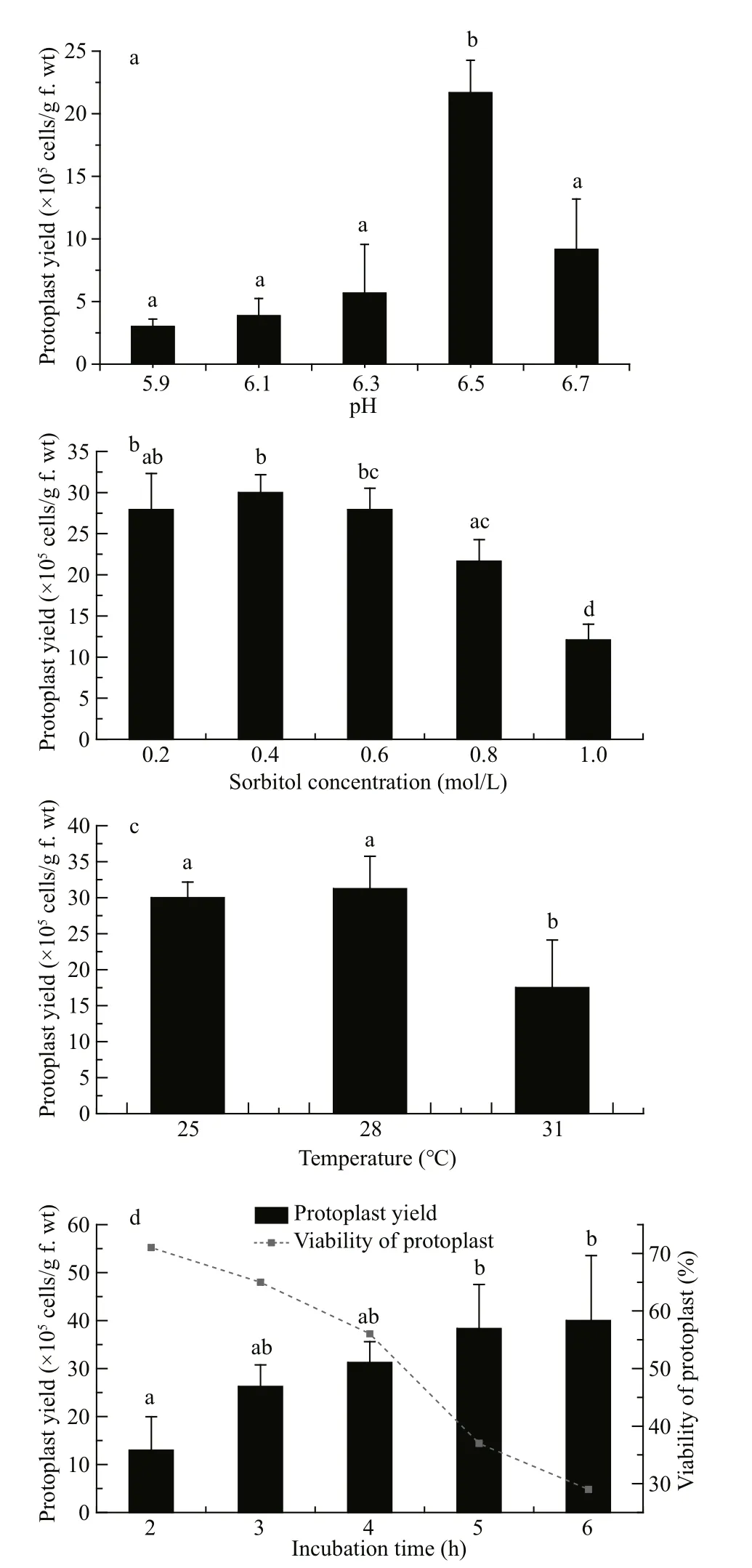
Fig.4 Effects of (a) different incubation pH, (b) sorbitol concentration, (c) incubation temperature, and (d)time on protoplast yield from G. bailiniae thalli
3.4 Optimization of incubation pH, sorbitol concentration and incubation temperature and time
pH 6.5 was found to be the optimal pH for protoplast yield ofG.bailiniae, with a significantly increased yield compared with the other pH values assessed (P<0.05) (Fig.4a).
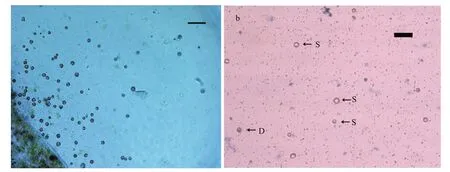
Fig.5 Protoplasts of G. bailin i ae
The protoplast yields were not significantly different with sorbitol treatments of 0.2, 0.4 and 0.6 mol/L (Fig.4b); the highest protoplast yield was obtained when the sorbitol concentration was 0.4 mol/L. The yield of protoplast decreased significantly (P<0.05) when the sorbitol concentration was >0.6 mol/L. Based on these results, a sorbitol concentration of 0.4 mol/L was considered optimal for protoplast yield.
Protoplast yields fromG.bailiniaewere not significantly different between the incubation temperatures of 25 and 28°C, and the optimal yield was obtained at 28°C. The protoplast yield decreased significantly (P<0.05) at 31°C (Fig.4c).
Protoplasts were released fromG.bailiniaeas early as 2 h after the start of incubation of the thalli in enzyme solution. Protoplast yields increased with incubation time (Fig.4d). However, the survival rate of the protoplasts decreased as the incubation time increased. As Fig.4d shows, the protoplast yield fromG.bailiniaeobtained after 4.0 h was(31.3±4.5)×105cells/g f.wt and the survival rate was 56%, the highest viable yield among the tested incubation times. Therefore, an incubation period of 4 h was considered optimal for viable protoplast yield.
3.5 Protoplast cell wall regeneration and cell division
Freshly isolated protoplasts fromG.bailiniaewere spherical with diameter from 7 to 35 μm (Fig.5a).Living protoplasts appeared yellow and dead protoplasts were dark blue when stained with Evans blue (Fig.5b). True protoplasts were red and undigested cell walls were green when they were stained with Fluorescent Brightener 28 and observed under UV light (Fig.6a). Regeneration of the cell wall began from one pole of the protoplast (Fig.6b). After being cultured for 48 h, the surface of most protoplasts was covered with regenerated cell wall and then the cell wall thickened. After 60 h of culture, the cell wall regeneration was almost complete (Fig.6c). We found that the first division of most protoplasts occurred on the third day after protoplasts were isolated. There were two division types for the first division of protoplasts fromG.bailiniae, asymmetric division and symmetric division (Fig.7a). In the former,protoplasts produced a small bud from one side of the cell, and in the latter, protoplasts divided into two cells from the cell middle. After the first division, the divided cells could develop into callus masses over several days of culture (Fig.7b, c).
4 DISCUSSION
Common sources of algal cell wall degrading enzymes are digestive gland juices of herbivorous marine invertebrates, culture filtrates of marine bacteria, and commercial preparations (Yan and Wang, 1993; Araki et al., 1998; Mussio and Rusig,2006; Reddy et al., 2008; Yeong et al., 2008; Gupta et al., 2013; Wang et al., 2014; Zhang et al., 2014).However, the enzyme activity of digestive gland juices from marine invertebrates varies with sampling time, sampling species and production batch of enzymes, and this material has highly deleterious effects on protoplast viability (Wang, 1994; Zhao et al., 2005). Commercial cell wall degrading enzyme preparations have sometimes been assessed for protoplast isolation fromGracilariaspecies. Yeong et al. (2008) and Huddy et al. (2013) obtained a large quantity of protoplasts fromGracilariachangiiandGracilariagracilisusing commercial cellulase,macerozyme and agarase. In our tests, protoplasts could not be isolated effectively fromGracilariopsis bailiniaeusing commercial agarase (TAKARA),cellulase and macerozyme R-10 (data not shown), but we obtained many protoplasts fromG.bailiniaewhen the commercial agarase was replaced by crude agarase solution fromMarinomonassp. YS-70. This might be because the commercial agarase is purer and could not completely degrade the complex amorphous matrix including sulfated polygalactans (agarcolloids or agars) in the cell wall ofG.bailiniae, resulting in non-release of protoplasts.
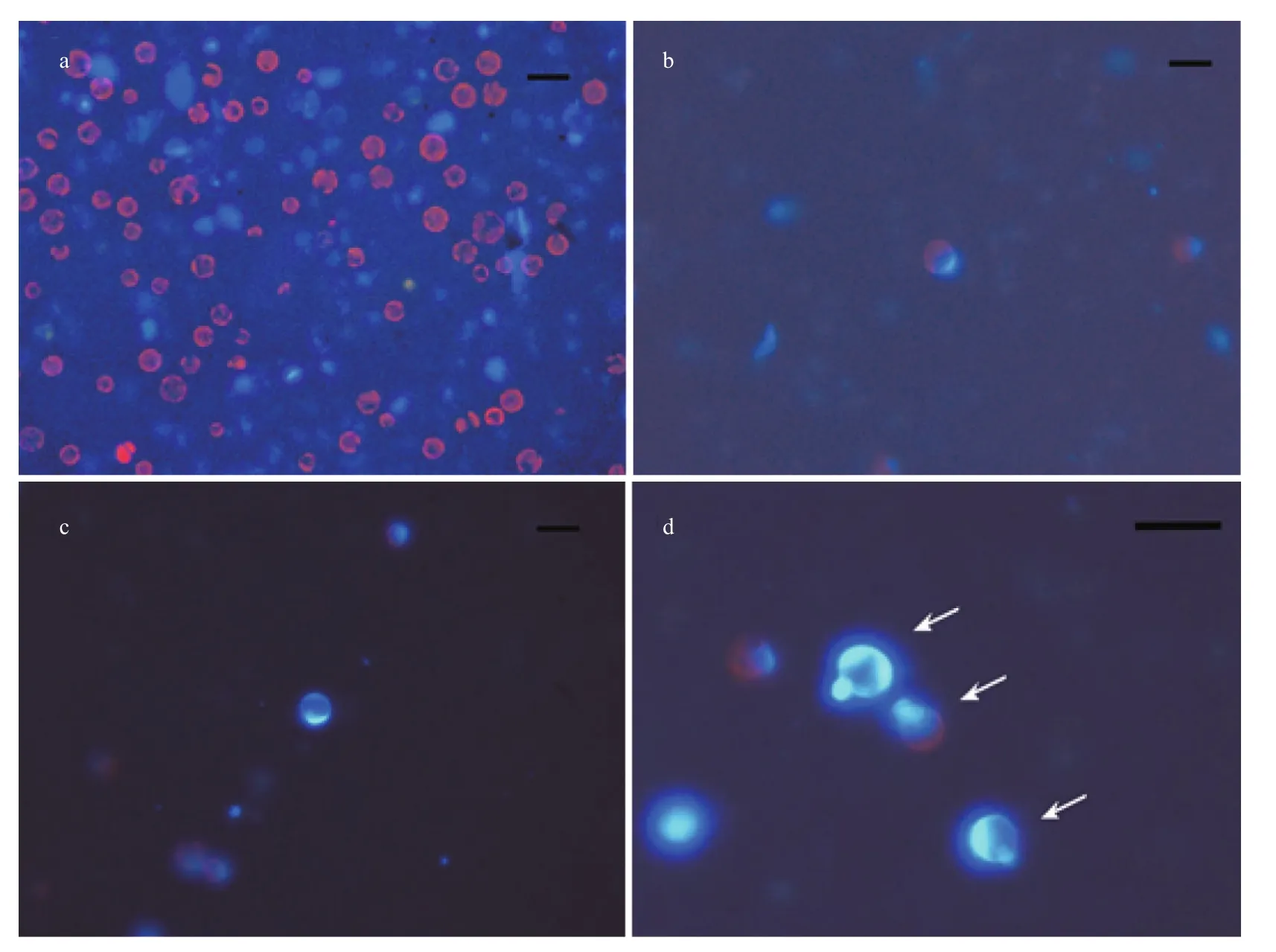
Fig.6 Cell wall regeneration of protoplasts from G. bailiniae
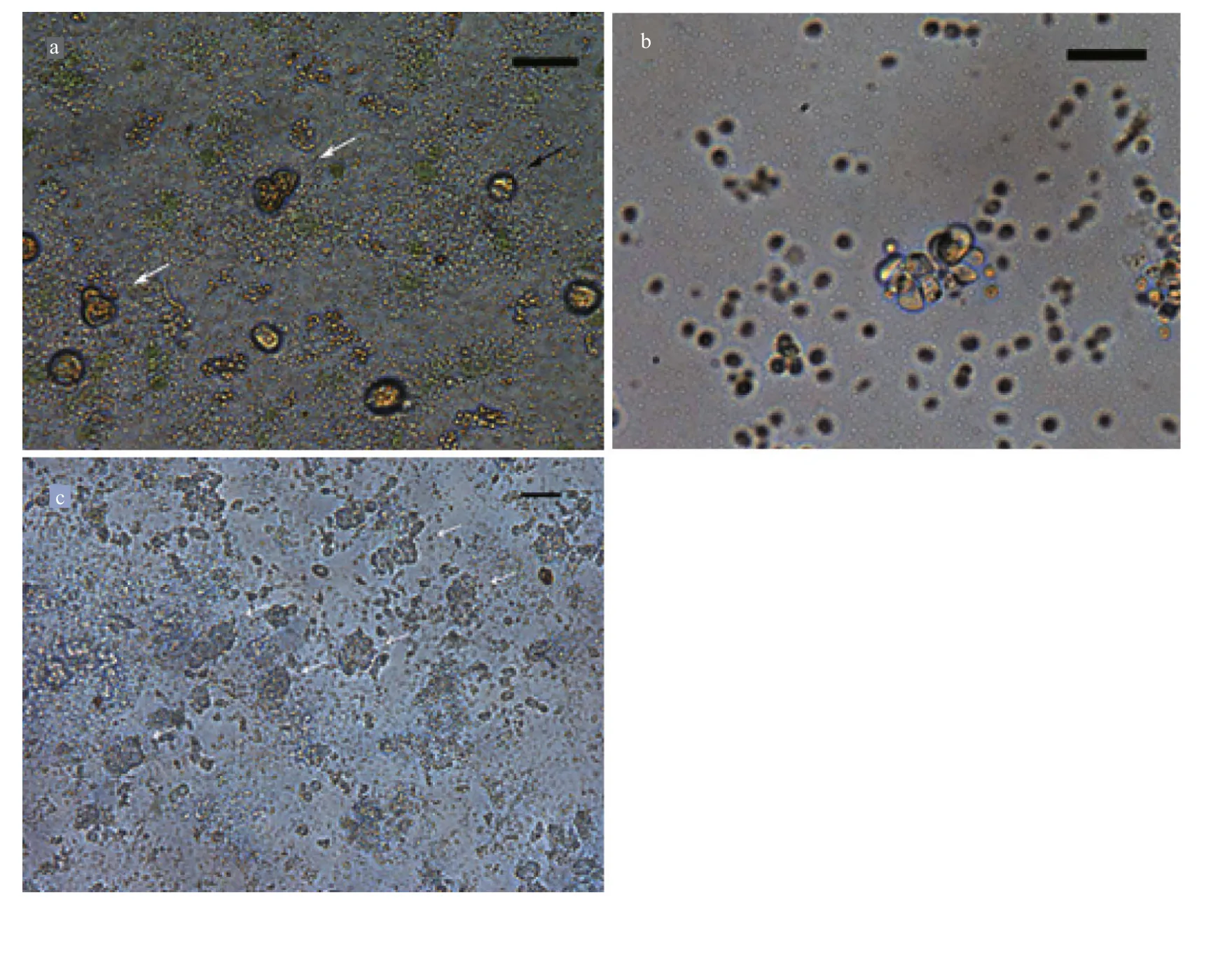
Fig.7 Protoplast regeneration of G. bailiniae
Studies have shown that red algae have a wide variety of matrix polysaccharides in their cell walls,which vary in amount depending on the species(Bellanger et al., 1990; Graham and Wilcox, 2000;Reddy et al., 2008). In our study, cellulase and agarase were necessary for protoplast isolation. We could not obtain a good protoplast yield fromG.bailiniaewith a single cell wall degrading enzyme or enzyme mixtures lacking agarase or cellulase, which suggests the cell wall ofG.bailiniaeis complex. Macerozyme R-10 presumably enhanced the protoplast yield because macerozyme has been shown to digest pectin and polygalactans in many studies of protoplast isolation from agarophyte species (Evans and Bravo,1983; Yeong et al., 2008; Gupta et al., 2011; Huddy et al., 2013). However, the commercial enzyme is partially purified and contains (a) toxic substance(s)(Inoue et al., 2011), so the optimization of cell wall degrading enzyme concentration was necessary. A combination of 2% w/v cellulase, 1% w/v macerozyme and 60% crude agarase prepared fromMarinomonassp. YS-70 resulted in the highest protoplast yield among the combinations and concentrations of enzymes we tested.
pH is an important parameter for protoplast isolation. In general, the pH used in published reports of algal protoplast isolation ranges between pH 5.8 and 7.0 (Cheney et al., 1986; Björk et al., 1990; Yan and Wang, 1993; Mussio and Rusig, 2006; Reddy et al., 2006; Yeong et al., 2008; Lafontaine et al., 2011).However, Gupta et al. (2011) reported an optimal pH of 7.5 for the isolation of protoplasts fromGracilaria verrucosaandGracilariadura. In our study, the optimal pH was 5.0 for commercial cellulase activity and 5.0 to 6.0 for macerozyme R-10, while the crude agarase fromMarinomonassp. YS-70 had its highest activity at pH 5.5–8.5 (Fig.1). Thus, weakly acidic or neutral solution (pH 5–7) could maintain relatively high activity of all three enzymes. The optimal pH of 6.5 for protoplast yield observed in this study is consistent with this conclusion.
Protoplast yield is influenced by many physicochemical factors, especially the constituents of the enzyme solution. In some previous studies of protoplast isolation from seaweed, CaCl2·2H2O, MES buffer, or both, were added into the enzyme solution(Dipakkore et al., 2005; Mussio and Rusig, 2006;Yeong et al., 2008; Huddy et al., 2013), but other studies did not use these reagents (Björk et al., 1990;Wang et al., 2014). In the present study, the protoplast yield was significantly enhanced by base-solution 3(deionized water containing 25 mmol/L MES-Tris buffer and 25 mmol/L CaCl2·2H2O). It is possible that the MES-Tris buffer maintained a stable pH for enzyme function and CaCl2·2H2O as a plasma membrane stabilizer protected the plasmalemma of the protoplasts (Wang, 1994). We found that a deionized water-based solution gave a higher protoplast yield than seawater based-solution. Similar results were also reported in studies of protoplast isolation fromUlva,EnteromorphaandMonostroma(Dipakkore et al., 2005). This phenomenon was possibly because ions in the seawater result in reduced activity of cell wall lytic enzymes (Dipakkore et al.,2005).
In this study, 0.4 mol/L sorbitol was found to be the optimal concentration for protoplast yield. Similarly,sorbitol was used as the osmotic stabilizer by Wang et al. (2014) in protoplast isolation fromGracilariopsis lemaneiformis, but the concentration of sorbitol in their study was 0.8 mol/L. Osmotic conditions help protoplasts to maintain their integrity after the cell walls are removed (Compton et al., 2000). Sugars and sugar alcohols, such as mannitol, sorbitol and glucose,are commonly used as osmotic stabilizers in protoplast isolation (Araki et al., 1998; Lafontaine et al., 2011;Wang et al., 2014; Zhang et al., 2014). Use of the inorganic osmoticum NaCl was also reported in protoplast isolation fromLaminariaspecies (Butler et al., 1989).
In our experiments, the optimal incubation temperature for protoplast yield fromG.bailiniaewas 28°C, close to the optimal growth temperature ofG.bailiniaein nature (Zhong et al., 2014). An optimal incubation temperature for protoplast isolation close to the optimal growth temperature of the algae was also observed for the tropical speciesGracilaria changiiand the temperate speciesGracilaria Verrucosa(Araki et al., 1998; Yeong et al., 2008;Gupta et al., 2011).
The protoplast yield increased, but the protoplast survival percentage decreased, when the incubation time increased during isolation. A similar result was found in the protoplast isolation fromKappaphycus alvarezii(Zhang et al., 2014). Optimal incubation times for protoplast isolation from many algae are between 2.5 and 3 h, for example 2.5 h forGracilaria verrucosa(Araki et al., 1998), and 3 h forGracilaria changii(Yeong et al., 2008),Gracilariagracilis(Huddy et al., 2013),MonostromanitidumandPorphyrayezoensis(Kito et al., 1998). Incubation times >6 h cause the cells to be overdigested, resulting in a significant reduction of the protoplast yield(Yeong et al., 2008).
Cell wall resynthesis of protoplasts was observed to start within 12 h (data not shown) after protoplast isolation fromG.bailiniae. Cell wall regeneration began at one pole of the protoplast. A similar pattern of cell wall deposition, beginning at a single pole of the protoplast, was also noted forGracilariagracilis(Huddy et al., 2013) andK.alvarezii(Zablackis et al.,1993). Two division types for the first division—asymmetric division and symmetric division—were first reported in the protoplast regeneration ofGracilariopsisgenera. After the first division, the divided cells can develop into callus-like masses. In the present study, many callus masses were obtained in the cultivation process, which might have potential for application in production of metabolites ofG.bailiniae. Meanwhile, some seaweeds, such asKappaphycusalvarezii(Reddy et al., 2003), have regenerated whole plants from their calli, so the callus masses obtained in our study might have the potential to produce seed material forG.bailiniae.
In previous reports, protoplasts ofGracilaria changiiandGracilariagracilisregenerated after 2 months of culture (Yeong et al., 2008; Huddy et al.,2015), and protoplasts ofGracilariopsislemaneiformisregenerated after 90 days of culture (Wang et al.,2014). However, we have been unable to regenerate protoplasts ofG.bailiniaeinto whole plants. The capacity for protoplast regeneration might be related to the growth conditions of the algal thalli. Using algae which are in good condition could improve the yield and the survival rate of protoplasts, and be good for protoplast regeneration (Björk et al., 1990; Wang et al., 2014). The thalli ofG.bailiniaeused in our study had been cultured for 1 month before protoplast isolation, so they might have been in poor condition and unsuitable for subsequent regeneration. In addition, the culture medium used in our study might not be suitable for protoplast regeneration ofG.bailiniae. Huddy et al. (2015) regeneratedGracilaria gracilisprotoplasts into whole plants in Provasoli’s enriched seawater (PES) medium. However, Yeong et al. (2008) obtained wholeGracilariachangiiplants from protoplasts only in MES medium but not PES medium. Moreover, the densities of protoplasts in our culture system were 5×104–1×105cells/mL, which might not be suitable for protoplast regeneration.Thus, further studies, especially to determine the optimum conditions for the growth of protoplasts into wholeG.bailiniaeplants, need to be conducted.
5 CONCLUSION
In this study, we established a protocol for protoplast isolation fromG.bailiniaeand investigated primary development of the protoplasts. This study thus provides a theoretical basis for seedling production ofG.bailiniae, and a protoplast isolation method for protoplast fusion and hybridization work.Furthermore, a large number of callus masses could be obtained in our study, which might have potential application in production ofG.bailiniaemetabolites.
6 ACKNOWLEDGEMENT
The authors thank Professor LIU Tao (Ocean University of China), Professor MEI Zhiping(Shantou University), Dr. WANG Hui (Shantou University) and Ms. WANG Zhongxia (Ocean University of China) for providing much advice and kind assistance.
猜你喜欢
西南科技大学学报(哲学社会科学版)(2022年1期)2022-11-21
宝藏(2022年2期)2022-07-30
鸭绿江(2020年21期)2020-10-14
农家之友(2020年2期)2020-05-19
青年生活(2019年6期)2019-09-10
汉字汉语研究(2019年1期)2019-05-21
黄河之声(2018年20期)2018-12-14
中学数学杂志(高中版)(2016年1期)2016-02-23
 Journal of Oceanology and Limnology2018年6期
Journal of Oceanology and Limnology2018年6期
- Journal of Oceanology and Limnology的其它文章
- Neuroanatomy and morphological diversity of brain cells from adult crayfish Cherax quadricarinatus*
- On the influence of season and salinity on the phenology of invertebrates in Australian saline lakes, with special reference to those of the Paroo in the semiarid inland
- Man-made plutonium radioisotopes in the salt lakes of the Crimean peninsula
- Cladophora mats in a Crimean hypersaline lake: structure,dynamics, and inhabiting animals
- Antimony speciation at the sediment-water interface of the Poyang Lake: response to seasonal variation*
- Seasonal variations of phosphorus species in the Tuohe River,China. Part I. Sediments*