
红松洼自然保护区草地群落物种多样性和土壤肥力与地上生物量的相关性研究
2019-01-09 10:28王瑶钱金平董建新
生态科学 2018年6期
王瑶, 钱金平, *, 董建新
红松洼自然保护区草地群落物种多样性和土壤肥力与地上生物量的相关性研究
王瑶1, 钱金平1, *, 董建新2
1. 河北师范大学资源与环境科学学院, 石家庄 050000 2. 河北民族师范学院生物与食品科学学院, 承德 067000
物种多样性与生物量之间的关系已经成为现代生态学研究的重要问题, 而土壤肥力也是影响生物量的重要因素。文章以红松洼自然保护区为例, 通过对红松洼自然保护区核心区和实验区植物的种类、数量、高度、盖度、物候期的实地采样调查, 以及对两区土壤的含水量、有机质含量、全氮含量、全磷含量的实地采样调查, 系统地分析了红松洼自然保护区植物物种多样性、土壤肥力与草地地上生物量三者之间的相关性。结果表明, 草地群落植物物种多样性与草地地上生物量呈显著正相关(<0.05), 植物物种多样性指数、均匀度指数与草地地上生物量也呈显著的正相关(<0.05), 土壤肥力中土壤水分、土壤全氮含量与草地地上生物量之间并无显著相关性(>0.05), 但是与土壤有机质含量呈显著的正相关性(<0.05), 与土壤全磷呈显著的负相关性(<0)。揭示可以通过提高植物物种多样性和土壤有机质含量来提高草地地上生物量。
物种多样性; 地上生物量; 土壤肥力; 草原生产力; 相关性
1 前言
草原生态系统占陆地总面积的20%左右, 是当前分布范围最广的植物类型[1]。草地植物群落的地上生物量是一种十分重要的草原生态系统功能[2]。出于科研目的和经济效益, 我们经常对草地的地上生物量进行调查统计, 从而更方便地维护草原植物群落的生态平衡。近年来, 草原受人为干扰退化严重, 草原生态安全问题已经引起学者的重视, 其中关于生物量与生物多样性的相关性研究层出不穷[3-6]。李中林等人认为, 过度放牧导致草地植被盖度减少, 生物多样性丧失, 草地群落的高度、密度和地上生物量均降低[7]; 郑伟等人认为, 地上生物量与物种多样性成单峰曲线关系, 即在中等生产力水平物种多样性最高的观点[8-9]; 白可喻等人认为, 草地生物多样性指数、均匀度指数与生物量之间呈极显著的正相关关系, 即生物多样性越高其生物量也越高, 而生物量与土壤水分相关性并不显著, 与土壤有机质和土壤全氮含量呈显著负相关[10]。还有一些国外学者认为, 物种多样性与生物量的关系表现为多种形式, 如单峰型、线性正相关、线性负相关、U型以及不相关关系[11]。
虽然学者们对物种多样性或土壤肥力与生物量的相关性做了大量研究, 但却主要集中在对其中两者之间的比较, 同时探讨三者间相关性的研究较少, 并且大都是针对不同区域大领域范围的研究, 在同一区域不同利用方式下三者间的相关性研究较少。对于红松洼国家级自然保护区来说, 由于草地利用方式不同, 导致物种多样性和土壤肥力不同, 同样也造成草地地上生物量发生显著变化。为了更好地保持草原生态系统稳定和可持续发展, 保证草地地上生物量能长久保持在较高水平, 文章在前者的基础上深入讨论, 实地采集了2015年、2016年红松洼地区物种多样性数据、地上生物量数据、土壤肥力数据, 构建三者的关联性, 并对保护区内核心区、实验区三者的关联性进行空间差异分析, 利用2015年和2016年的数据作对比, 分析不同利用方式下三者关系的时间演变格局, 在时间和空间上分析红松洼自然保护区物种多样性、土壤肥力与地上生物量之间的作用关系, 建立综合相关性评价体系, 尝试从物种多样性和土壤肥力方面维护草地生态系统平衡,从而为保护各级自然保护区提供科学的依据。
2 材料与方法
2.1 研究区自然概况
红松洼自然保护区位于河北省围场满族蒙古族自治县境内, 地理坐标为N42°10′—N42°20′, E117°18′—117°35′, 总面积7300公顷, 是清代皇家猎苑木兰围场的一部分。1994年经河北省人民政府批准建立, 并于1998年晋升为国家级自然保护区, 是一个以亚高山草甸为主要植被类型的自然保护区, 属于中温型大陆季风气候, 冬季寒冷漫长, 夏季凉爽短暂, 昼夜温差大, 积雪期长达7个月, 平均年降水量为450 mm—550 mm。主要以山地为主, 海拔在1600 m—1800 m之间。保护区内有种子植物595 种, 药用植物81 种, 观赏植物60 多种, 物种多样性丰富, 植被覆盖度达到67.76%。土壤多为草甸土和山地黑土, 命名为红松洼黑土, 土壤肥力肥沃, 腐殖质含量为6.54%, 氮含量为0.294%, 磷含量为0.108%, 土壤pH值为6.5—6.9, 剖面30 cm以下有中量白色二氧化硅粉末。1 m以下出现大量锈纹锈斑(图1)。
2.2 研究方法
2.2.1 群落调查和取样方法
红松洼自然保护区核心区实施了围栏封育的政策, 导致各区植物生长状态发生改变。因此在红松洼自然保护区的核心区和实验区各设置一个正南正北方向100 m×100 m的大样方, 样地选在较为平坦且无灌木的地方, 保持两地的光照条件和地形情况相同, 核心区的大样方记做H1, 实验区的大样方记做H2, 用GPS机定位样地坐标, 并记录。之后在每个大样方中设置25个1 m×1 m的小样方, 样方采取从东南角开始每隔20 m选取一个小样方的平均取样法, 并按Z字形排列, 以保证取样的随机性和记录的方便性。在核心区将每个小样方按照顺序分别记做H1-1—H1-25, 在实验区记做H2-1—H2-25。在植物生长高峰期的7月和8月期间[12], 将小样方内地上植物齐地割下, 测定群落的种类组成和其特征值(数量、高度、盖度、干重), 测量样方中各种植物的地上生物量, 并记录位置信息。

图1 红松洼国家级自然保护区遥感影像图
土壤取样是在小样地中, 用土钻钻取每个小样方地下0—30 cm深土样, 并按照位置进行标记, 将土壤标号后带回实验室进行检测。过筛后称量湿重, 而后烘干(烘干采用放入恒温80 ℃的烘干机中进行烘干), 测量干重[12]。烘干后测定土壤的基本养分, 测量土壤的全磷、全氮、有机质的含量。土壤全磷用钼锑抗比色法[12], 土壤的全氮含量采用凯氏定氮法(Foss kjeltec 2300)测定[12], 土壤有机质的测定选择重铬酸钾法[13]。
2.2.2 物种多样性测度测定方法
根据2014年国家环保部发布的《生物多样性观测技术导则—陆生维管植物》环境保护标准公告, 该研究利用多样性的测度方法。多样性是指在栖息地或群落中的物种多样性, 用以测度群落内的物种多样性。测度多样性采用物种丰富度(物种数量)、辛普森(Simpson)指数、香农—维纳(Shannon- wiener)指数、皮洛(Pielou)均匀度指数和Margale指数[14]。
物种丰富度指数:=(此时为总物种数)
Simpson多样性指数():
=1−∑(/)²
Shannon-wiener指数():
=-∑L∕ln(/)
Margalef指数():
=(-1)/ln
Pielou均匀度指数():
=/ln
式中,为Shannon-wiener指数,为群落中的物种总数目,为观察到的个体总数,为第个物种所在样方的物种数。
Simpson指数是测定物种多样性的一个有力指标, 可以明确表示出物种的丰富度和物种的均匀度; Shannon指数用来表示群落多样性的高低, 从而对比计算出数据, 得出结论; Margelaf指数用来表示物种丰富度; Pielou均匀度指数用来表示物种分布的均匀程度。
2.2.3 分析方法
运用Excel2007对所有的数据进行输入和简单运算处理, 对于复杂的数据处理如双变量相关分析, 则采用spss16.0进行数据分析和统计。利用Pearson进行相关性分析, 并通过双尾进行显著性检验。
3 结果与分析
3.1 不同草类对草地生物量的贡献
我们可以将草地按功能分类, 分别为牧草、杂草和药用草。从整体上看, 三种草类物种数量核心区均多于实验区, 对样地内草类进行更细致的统计分析, 得出2015年核心区牧草为12 种、杂草15 种、药用草28 种; 实验区牧草11 种、杂草9 种、药用草22 种; 2016年核心区牧草10 种、杂草14 种、药用草19 种;实验区牧草7 种、杂草12 种、药用草18种。得出结论:核心区植物种类数量为药用草>杂草>牧草;实验区植物种类数量在2015年为药用草>牧草>杂草, 2016年为药用草>杂草>牧草。药用草无论在试验区还是在核心区种类都最多。
在空间位置上对不同草类的地上生物量做比较, 总体来看, 地上生物量核心区均大于实验区, 2015年实验区比核心区低21.2%, 2016年实验区比核心区低68%, 而两年核心区地上总生物量变化不明显, 说明实验区的草地可能处于加速退化阶段。牧草地上生物量在2015年实验区比核心区低64.8%, 在2016年实验区比核心区低53%, 总体上仍表现为实验区低于核心区;杂草类地上生物量在2015年两区的差距不大, 实验区较核心区低4%, 而在2016年实验区较核心区低75.3%, 可以看出实验区杂草类的退化情况严重;药用植物地上生物量在2015年实验区较核心区低55%、在2016年实验区较核心区低2.3%(表1)。在该区, 人类干扰主要包括公路建设、旅游开发、风电开发建设、草地围栏、放牧(全年放牧、冷季放牧、暖季放牧、春秋放牧)、采集等活动。上述人类干扰在一定程度上均会对草地种类和生物量产生影响, 其中对牧草影响最大的是放牧, 大量牦牛、羊的选择性进食导致实验区草地地上生物量降低;对杂草影响最大的是旅游和风电, 两者会对土壤造成破坏, 降低土壤养分, 杂草无法汲取足够的养分引起退化; 对药用植物影响最大的是草地围栏和采集, 由于并未对实验区进行围栏封育, 当地居民、游客对药用植物的选择性采摘, 降低了实验区药用植物的数量, 而核心区药用植物数量的急剧下降是由于种间竞争引起的, 药用植物竞争力较弱, 逐渐被牧草替代。
再对数据进行时间上的对比。在核心区, 2016年比2015年的各类草种数量都有所减少, 总干重也较2015年有所减少, 总地上生物量差异虽不太明显, 但是杂草和药用草类生物量减少, 而牧草类生物量较上年上升了227%(表1)。牧草种类虽然减少但是地上生物量却呈上升趋势, 主要是由于物种间的竞争效应引起的, 优势种逐渐代替劣势种, 说明牧草已经成为核心区的优势种。在实验区, 除牧草种以外, 2016年比2015年其他草类种数都有所下降, 其中杂草类数量和总干重差异最为显著, 是由于旅游、科研、风电开发等活动造成的破坏比放牧造成的破坏更为严重, 造成实验区牧草种逐渐替代杂草和药用植物, 处于优势地位, 从总体上造成草地严重退化的后果。得出以下结论:由于实验区允许放牧、采摘、科研、旅游、风电等活动进行, 牛羊对优良牧草的选择性进食, 人类对有利用价值的药用草类有目的性采摘等, 都是造成实验区牧草地上生物量低于核心区的根本原因。而核心区由于围栏封育, 对草原保护良好, 牧草的地上生物量不断提高, 又由于在一定范围内物种的种间竞争, 造成牧草种逐步成为优势种, 而杂草和药用植物成为劣势种, 处于退化阶段。
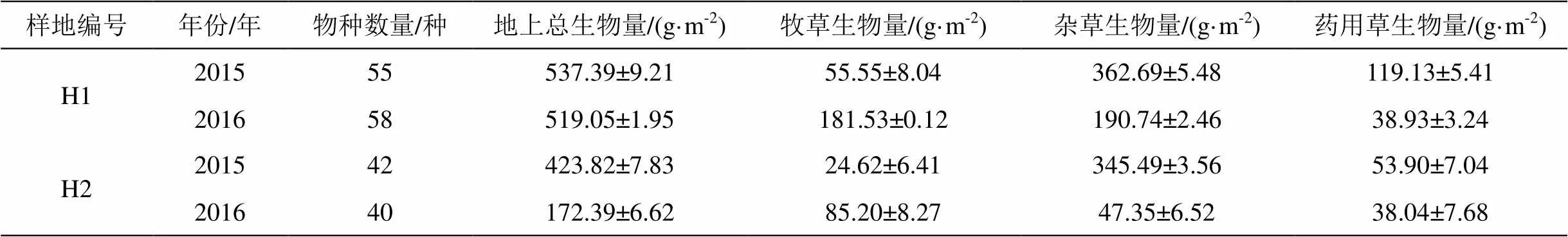
表1 不同利用方式下的地上生物量

图2 地上生物量变化
综上所述, 在核心区对地上生物量贡献最多的物种是杂草类, 但是由于围栏封育政策的实施, 核心区牧草种地上生物量呈现出逐年递增的局面, 在未来牧草种将成为核心区的优势种, 提高草地地上生物量。在实验区, 杂草类和药用植物类都受到人类活动的影响, 地上生物量处于快速退化的阶段, 而牧草种由于牛羊的进食反而增加了它的生长速度, 整体来看变化不明显, 处于轻微退化阶段。
3.2 草地植物物种多样性和土壤肥力
实验得出, 两个样方中核心区的植物物种多样性指数和均匀度指数均高于实验区(表2), 并且Simpson多样性指数和Shannon-wiener多样性指数与Pielou均匀度指数均呈正相关(<0.05), Margalef指数与Pielou均匀度指数相关性不显著(>0.05)(表4), 说明在植物物种多样性比较高的核心区, 物种均匀度也较高, 两者呈现出显著正相关性(<0.05)。2015年核心区植被覆盖率为66%, 实验区为45%;2016年核心区植被覆盖率为65%, 实验区为32%。核心区的植被覆盖率显著高于实验区, 也表明核心区的植物物种多样性高于实验区。对两年数据进行分析, 该结果主要是由人为干扰引起的。核心区草地严格实施围栏封育策略, 实施禁牧、禁入、不允许开发风电和旅游项目等措施, 使核心区草地受到良好的保护;在实验区由于超载放牧、人为践踏、车轮碾压、实验科研、开发风电旅游等活动, 造成实验区物种多样性减少, 其多样性指数、均匀度指数均显著低于核心区, 造成两地的植被覆盖率差异显著。
本研究中, 土壤肥力主要是指土壤有机质含量、土壤含水量、土壤全氮全磷含量。在红松洼自然保护区, 土壤肥力除了土壤全氮含量在2015年是H2>H1以外, 其余均为H1>H2。分析其原因, 土壤全氮主要是由植物根部的根瘤菌将空气中的氮素固定到土壤中;土壤磷肥主要来源于生物的排泄物和生物遗体, 核心区物种数量多, 植株数量大, 动物排泄物和动植物在秋冬季节遗体数量均较多, 土壤中磷肥含量也随之增加。由于核心区长期围栏封育, 物种多样性较高, 草类种类增多, 个体数量增加, 秋季枯物数量增加, 导致土壤有机质含量增加。总体来说, 两年土壤肥力H1>H2。
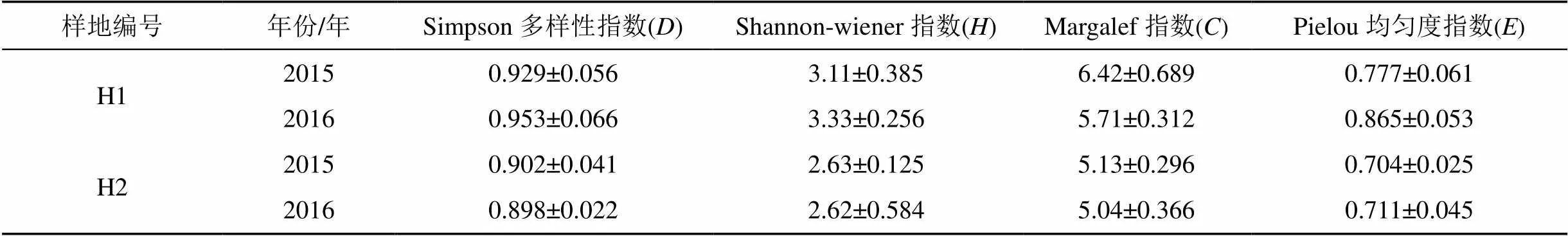
表2 多样性指数计算

表3 土壤肥力含量
3.3 草地物种多样性与地上生物量之间的关系
草地地上生物量, 2015年核心区多于实验区1133.6388 kg·hm-2, 实验区占核心区的78.9%; 2016年核心区高于实验区3466.5536 kg·hm-2, 实验区仅占核心区的33.2%。而核心区两年的地上生物量基本没有变化, 保持在5200 kg·hm-2, 属于较高生产力;实验区的地上生物量平均值为2900 kg·hm-2, 地上生物量相对较低, 且正处于退化阶段(表2)。数据对比说明, 实验区的草场退化十分严重, 是亟待解决的草原生态问题。
对调查样地的草地植物物种多样性和草地地上生物量做Pearson相关性分析, 可以看出草地地上生物量与Simpson多样性指数呈显著正相关(<0.05), 草地地上生物量与Pielou均匀度指数呈显著正相关(<0.05), 说明物种多样性越丰富, 物种分布越均匀, 抵御外界干扰的能力越强, 草原地上生物量越大, 草原生态系统越稳定, 两者呈线性正相关(表4)。
3.4 土壤肥力与草地地上生物量之间的关系
实验对土壤含水量、有机质含量、土壤全氮、土壤全磷和地上生物量做Pearson相关性分析, 可以看出土壤有机质与草原地上生物量呈显著正相关(<0.05), 与土壤全氮、土壤含水量相关性不显著(>0.05), 与土壤全磷呈显著的负相关(<0), 全氮含量与全磷含量呈现出负的相关性(<0)(表4)。说明地上生物量随有机质含量的增加而增加, 随土壤全磷含量的增加而减少, 与土壤全氮、土壤含水量无显著相关性。
对上述结果进行分析, 在土壤有机质含量方面, 核心区与实验区相比, 前者植物物种多样性较大, 植物数量较多并且土壤有机质99%来源于植被的腐殖质, 秋冬季植物腐殖质较多, 包括各种植物残体、微生物、动物残体都会增加土壤有机质含量从而增强土壤肥力;而在实验区, 植被覆盖度较低, 裸露的地表由于没有草层覆盖导致土壤肥力逐年降低, 土层破坏严重, 物种种类较为单一, 动植物残体形成的土壤有机质含量较少, 土壤肥力下降, 草地地上生物量也随之下降, 从而形成一个恶性循环系统, 所以土壤有机质含量与草地地上生物量呈显著正相关。在土壤含水量方面, 本实验结果与一些学者研究相符[11], 我认为这一结果是片面的, 该结论在同一地块成立, 在不同地块应相反。在红松洼自然保护区H1、H2两地地理位置靠近, 土壤含水量差别不大, 属于同一地块, 所以在本研究中土壤含水量与草地地上生物量之间并无相关性。在土壤全氮含量方面, 虽然核心区固氮作用较大, 但是植物生长消耗的氮素也多, 因此与草地地上生物量之间的相关性不显著。在土壤全磷含量方面, 物种对于磷肥的吸收越多, 物种多样性越丰富, 反而导致土壤含磷量减少, 形成显著负相关性(<0)。
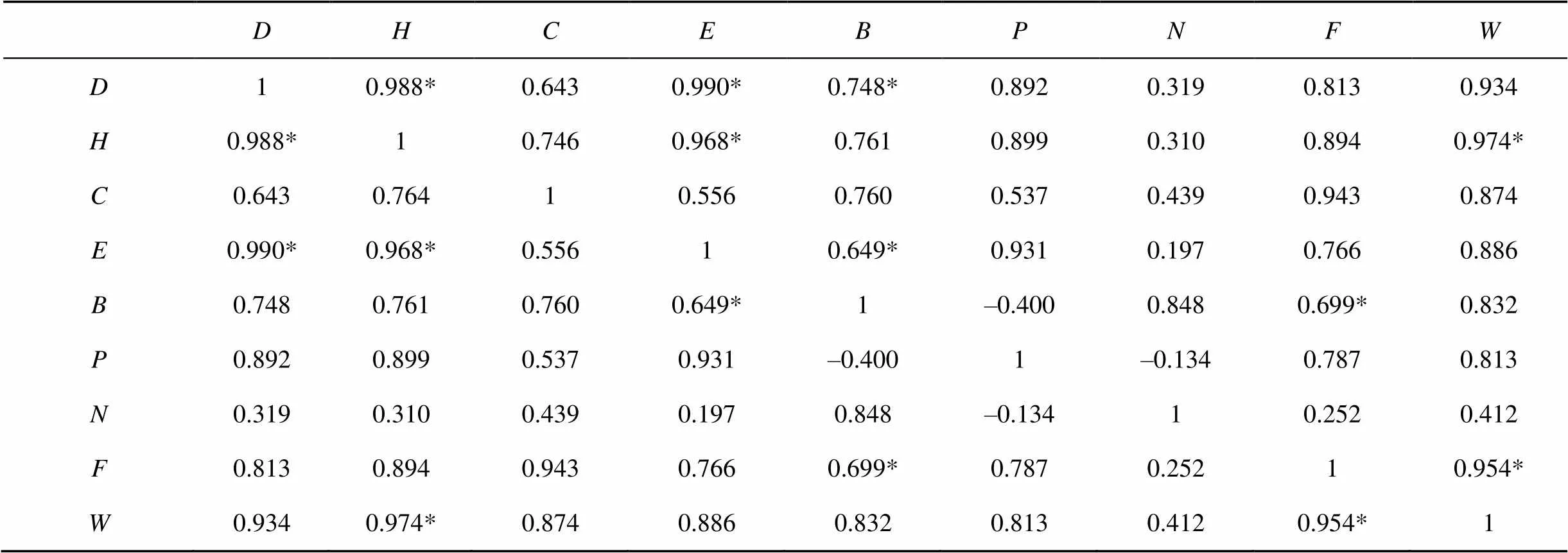
表4 地物种多样性与土壤肥力和草地地上生物量的皮尔森(Pearson)相关性分析
注:: Simpson多样性指数;: Shannon-wiener指数;: Margalef指数;: Pielou均匀度指数;: 地上生物量;: 土壤全磷;: 土壤全氮;: 土壤有机质;: 土壤含水量; *<0.05。
4 讨论
经过对物种重要值分析, 计算出核心区优势种为: 宽叶蒿()、细叶苔草()、裂叶蒿()、硬质早熟禾()、直穗鹅观草()、山岩黄芪()、地榆()、小黄花菜()、无芒雀麦();实验区优势种为: 细叶苔草()、硬质早熟禾()、珠芽蓼()、裂叶蒿()、宽叶蒿()、地榆()、越桔柳()、毛茛()、柳叶蒿()。其中, 虽然杂草对地上生物量贡献最大, 但是杂草却处于退化阶段, 未来将会被牧草替代。但在整体上并不影响物种多样性与地上生物量之间的正相关性。实验区生态环境遭受破坏严重, 土壤肥力退化, 造成物种多样性减少, 草原地上生物量降低。
对植物物种多样性、土壤肥力与地上生物量的相关性分析研究, 不仅从空间上对比两块样地利用方式不同所造成的差异, 还从时间上讨论了两地植物物种多样性与草原地上生物量的发展趋势, 从三维空间分析了二者之间的相关性。本研究表明, 物种多样性与地上生物量之间存在显著正相关关系(<0.05), 说明物种多样性与植物分布的均匀度成正比, 与王长亭(2004)[12]、Aarssen L.W(2004)[22]等人的研究结果一致, 而与郑伟、董全民(2013)[9]等人研究生物量与物种多样性成单峰曲线关系的结果不一致。对土壤肥力与草地地上生物量进行多方位分析, 实验结果表明, 草地地上生物量与土壤有机质含量呈显著正相关(<0.05), 与李亚娟(2016)[25]等人的地上生物量与土壤有机质含量呈显著正相关的结果一致; 与土壤全磷呈显著负相关(<0), 由于植物对磷肥的吸收作用, 随着植物数量增多土壤磷肥含量减少。这与左小安(2007)[18]等人的研究结果相违背, 草地地上生物量与土壤肥力之间并不是简单的线性关系。
本研究结论只考虑了影响生物量的植物物种多样性、土壤肥力两个因素。除此以外, 草原地上生物量还与其他诸多因素如气候、人类干扰等密切相关, 还有待进一步研究。
5 结论
通过实验研究得出以下结论: 1、杂草类对草地地上生物量的贡献值最大, 但目前处于退化阶段; 其次是牧草的地上生物量, 并且牧草的地上生物量处于上升阶段;药用植物对草地地上生物量的贡献最小。2、核心区的优势种有宽叶蒿、细叶苔草、裂叶蒿、硬质早熟禾、直穗鹅观草、山岩黄芪、地榆、小黄花菜、无芒雀麦;实验区优势种为细叶苔草、硬质早熟禾、珠芽蓼、裂叶蒿、宽叶蒿、地榆、越桔柳、毛茛、柳叶蒿。两区优势种稍有不同, 但数量差异显著。3、Simpson多样性指数、Shannon-wiener指数、Margalef指数、Pielou均匀度指数均是核心区>实验区。土壤有机质含量核心区>实验区, 并且核心区呈逐年减少, 实验区呈逐年增多的趋势;土壤全氮含量、土壤全磷含量均为核心区>实验区, 但两者与地上生物量关系不大。4、Simpson多样性指数、Shannon-wiener指数和Pielou均匀度指数均呈现显著正相关(<0.05); Simpson多样性指数和Pielou均匀度指数均与地上生物量成显著正相关(<0.05)。说明草地地上生物量受草原植物物种多样性与植物物种分布均匀度的直接影响, 呈现正相关关系。5、草原地上生物量与土壤有机质呈显著正相关(<0.05), 与土壤全磷呈显著负相关(<0), 与土壤含水量和土壤全氮相关性不显著(>0.05), 表明土壤有机质含量增加有利于草类植物生长, 土壤磷肥与草原地上生物量成逆向增长模式, 本研究中草原地上生物量与土壤含水量和全氮含量无明显关系。因此, 在一定范围内, 相同气候条件下, 植物物种多样性越高、土壤有机质含量越高, 草地地上生物量越高。我们在提高生物量时应着重从物种多样性和土壤有机质方面入手, 通过人工干扰(例如禁牧、规划放牧、种植牧草、人工施肥等措施), 在物种较稀少地区人工提高物种丰富度, 通过最大化植物物种多样性来提高草地地上生物量, 或者在植被植物多样性较弱地区实施人工增加土壤有机质肥, 从而达到提升产量的效果。提高草原生态平衡能力, 还要注意防止其他物种入侵引起的影响。综上所述, 只有植物均匀分布且植物物种多样性、土壤有机质均保持在较高水平, 才能保持草地地上生物量的较高水平。
[1] 金樑, 孙莉, 王强, 等. AM真菌在草原生态系统中的功能[J]. 生态学报, 2016, 36(3): 873–882.
[2] 石红霄, 侯向阳, 师尚礼, 等. 高山嵩草草甸初级生产力、多样性与土壤因子的关系[J]. 草业学报, 2015, 25(10): 40–47.
[3] BERHE M B K. Training reduced subjectivity of comparative yield method of estimation of grassland biomass[J]. Grass Forage Science, 2016, 71(3): 482–489.
[4] HEGAZY A K, EL M A , HOSNI H A. Vegetation, species diversity and floristic relations along an altitudinal gradient in south-west Saudi Arabia[J].Journalof Arid Environments, 1998, 38(1): 3–13.
[5] SCHALLER J, ROSCHER C, HILLEBRAND H , etal. Plant diversity and functional groups affect Si and Ca pools in aboveground biomass of grassland systems[J]. Oecologia, 2016, 182(1): 1–10.
[6] 刘哲, 李奇, 陈懂懂, 等. 青藏高原高寒草甸物种多样性的海拔梯度分布格局及对地上生物量的影响[J]. 生物多样性, 2015, 23(4): 451–462.
[7] 李中林, 秦卫华, 周守标, 等. 短期围栏封育对红松洼自然保护区群落数量特征的影响[J]. 草地学报, 2015, 23(1): 21–26.
[8] MITTELBACH G G, STEINER C F, SCHEINER S M, et al. What is the obversed relationship between species richness and productivity?[J]. Ecology, 2001, 82(9): 2381– 2396.
[9] 郑伟, 董全民, 李世雄, 等. 放牧强度对环青海湖高寒草原群落物种多样性和生产力的影响[J]. 草地学报, 2012, 20(6): 1033–1038.
[10] 白可喻, 戎郁萍, 杨云卉, 等. 北方农牧交错带草地生物多样性与草地生产力和土壤状况的关系[J]. 生态学杂志, 2013, 32(1): 22–26.
[11] WAIDE R B, WILLING M R, STENINER C F, et al. The relationship between primary productivity and species richness[J]. Annual Review of Ecology and Systematics, 1999, 30(3): 257–300.
[12] 王长庭, 龙瑞军, 曹光敏, 等. 高寒草甸不同类型草地土壤养分与物种多样性——生产力关系[J]. 土壤通报, 2008, 39(1): 1–8.
[13] 董全民, 赵新全, 马玉寿. 放牧强度对高寒混播草地土壤养分含量的影响[J]. 土壤通报, 2008, 39(3): 553–557.
[14] 生物多样性观测技术导则-陆生维管植物[S], 国家环保部, 2014.
[15] 谢应忠. 生物多样性的生态学意义及其基本测度方法[J]. 农业科学研究, 1998, 19(3): 13–20.
[16] 孙凡洲, 于仁成, 徐子钧, 等. 应用excel软件计算生物多样性指数[J]. 海洋科学, 2012, 36(4): 57–62.
[17] 刘海峰, 李连芳. 红松洼自然保护区草甸群落结构与多样性对管理模式的响应[J]. 生态学杂志, 2012, 31(5): 1082–1087.
[18] 左小安, 赵学勇, 赵哈林, 等. 科尔沁沙质草地群落物种多样性、生产力与土壤特性的关系[J]. 环境科学, 2007, 28(5): 945–951.
[19] JONATHAN L, NICOLE M G. Patterns of plant diversity and environmental relationships across three riparian corridors [J]. Forest Ecology and management, 2005, 20(4): 267–278.
[20] 景美玲, 马玉寿, 李世雄, 等. 氮肥对大通河上游退化草地的影响[J]. 草地学报, 2016, 24(3): 518–523.
[21] 贾文雄, 王洁, 张禹舜, 等. 祁连山南坡灌丛草甸生物量变化与水热因子的关系研究[J]. 地理科学, 2016(8): 1243–1251.
[22] AARSSEN L W. Interpreting covariaton in species richness and productivity in terrestrial vegetation: make sense of causation and correlations at multiple scales [J]. Folia Geobotanica, 2004, 39: 385–403.
[23] 字洪标, 阿的鲁骥, 刘敏, 等. 高寒草甸不同类型草地群落特征及优势种植物生态位差异[J]. 应用与环境生物学报, 2016 (4): 546–554.
[24] 刘玉, 常小峰, 田福平, 等. 放牧对草地群落与土壤特征的影响[J]. 西北植物学报, 2016, 36 (12): 2524–2532.
[25] 李亚娟, 曹广民, 龙瑞军, 等. 三江源区土地利用方式对草地植物生物量及土壤特性的影响[J].草地学报, 2016, 24 (3): 524–529.
[26] 刘艳, 宋同清, 蔡德所, 等. 喀斯特峰丛洼地不同土地利用方式土壤肥力特征[J]. 应用生态学报, 2014, 25(6): 1561–1568.
[27] 李连方, 王培, 王警龙, 等. 红松洼草地自然保护区种子植物区系的研究[J]. 中国草地学报, 1999 (5): 35–41.
[28] 董婷婷, 宝音陶格涛. 羊草人工草地生物多样性与生产力关系研究[J]. 中国草地学报, 2012, 34(3): 34–41.
[29] 张涛, 陈晓鹏, 赵景学, 等. 碳、氮添加对高寒草甸植物群落物种多样性和生物量的影响[J]. 生态环境学报, 2015, 24(10): 1604–1610.
The grass species diversity and soil fertility research of biomass on the ground of Hongsongwa Nature Reserve
WANG Yao1, QIAN Jinping1, *, DONG Jianxin2
1. Resources and EnvironmentalScience of Hebei Normal University, Shijiazhuang 050000, China 2.College of Biology and Food ScienceofHebei Normal University, Chengde 067000, China
The relationship between biodiversity and biomass has become an important problem in modern ecology research. Plant growth and development are related with soil factors. We took Hongsongwa Nature Reserve as an example, to investigatethe relationship by analyzing the plant species, quantity, height, and coverage, as well as the parameters including soil water content, organic matter content, total nitrogen, and total phosphorus content. The results showed that species diversity and above biomass of grassland were significantly and positively correlated (< 0.05); the grass species diversity index and evennessindex and above biomass of grassland also were significantly and positively correlated (< 0.05). Soilmoisture and soil total nitrogen content were not related with the above biomass of grassland (> 0.05), positively related with the soil organic matter content (< 0.05), but negatively related with soil total phosphorus (< 0). The biomass of grassland can be increased by increasing species diversity and soil organic mattercontent.
species diversity; ground biomass; soil fertility; grassland productivity; correlation
10.14108/j.cnki.1008-8873.2018.06.004
Q178
A
1008-8873(2018)06-030-08
2017-01-20;
2018-01-09
国家环保部关于红松洼自然保护区生物多样性调查(005049); 河北省自然科学基金项目: 太行山森林植被变化水文生态效应研究(D2013205187); 河北省承德市财政局资助项目(CZ3013003)
王瑶(1993—) , 女, 河北邯郸人, 硕士, 主要从事草原生态环境、资源与环境影响评价研究, E-mail:793496049@qq.com
钱金平, 男, 硕士, 教授, 主要从事区域资源开发利用规划生态环境保护的研究, E-mail: 793496049@qq.com
王瑶, 钱金平, 董建新. 红松洼自然保护区草地群落物种多样性和土壤肥力与地上生物量的相关性研究[J]. 生态科学, 2018, 37(6): 30-37.
WANG Yao, QIAN Jinping, DONG Jianxin. The grass species diversity and soil fertility research of biomass on the ground of Hongsongwa Nature Reserve[J]. Ecological Science, 2018, 37(6): 30-37.
猜你喜欢
西藏农业科技(2018年4期)2018-04-25
青海政报(2017年18期)2018-01-31
种业导刊(2017年5期)2017-06-27
种业导刊(2017年4期)2017-06-19
种业导刊(2017年3期)2017-06-19
投资北京(2017年1期)2017-02-13
基础教育参考(2015年5期)2015-06-01
茶叶(2014年4期)2014-02-27
城市道桥与防洪(2013年8期)2013-03-11
植物营养与肥料学报(2011年3期)2011-10-24
