
水稻恶苗病菌对咪鲜胺的抗性初探
2019-01-25 05:25赵渊高松刘连盟黎起秦黄世文
浙江农业科学 2019年1期
赵渊,高松,刘连盟,黎起秦,黄世文*
(1.广西大学 农学院,广西 南宁 530003; 2.中国水稻研究所 水稻生物学国家重点实验室,浙江 杭州 310006;3.河南农业大学 植物保护学院,河南 郑州 450002)
水稻恶苗病是由藤仓镰孢菌(Fusariumfujikuroi)侵染引起的一种真菌病害[1-3]。药剂种子处理是目前唯一可以依靠的防治该病的有效手段[4-6]。咪鲜胺是防控水稻恶苗病的主要药剂,在生产上应用广泛。近年来,由于抗性问题严重,咪鲜胺的药效降低,用量逐年加大[7-9]。
20世纪70—80年代,国内外接连报道发现苯菌灵抗性菌株,此时距苯并咪唑类杀菌剂大规模用于水稻恶苗病防治仅仅十几年时间[10]。据日本有关报道,20世纪80年代末到90年代初,对日本8个地区9个县采集的病样进行抗性测定,发现恶苗病菌对苯并咪唑类药物的抗性已达很高水平,其中,最低抑菌浓度(MIC)≥100 mg·kg-1的菌株占83.4%[11-12]。在我国,镇江农科所于1990—1992年连续3 a分别对全国4省市共16个县采集的菌株进行苯并咪唑类药物抗性测定,结果表明,MIC≥100 mg·kg-1菌株占所采集菌株的65.9%,抗性菌株已成为这些地区的优势种群[13]。目前,我国水稻恶苗病菌对多菌灵的抗性问题已经非常普遍,局部地区的抗性水平已达5 000倍以上[14]。
20世纪90年代,日本几家大型农药生产企业均分离到了对咪唑类农药不敏感的菌株,这距咪鲜胺的大面积推广应用尚不足十年[15]。赵志华等[16]研究表明,水稻恶苗病菌对咪鲜胺具有潜在的抗性风险;刘永锋等[17]研究表明,江苏省的田间恶苗病菌对咪鲜胺已有一定的抗性;陈夕军等[18]分离到了8株抗咪鲜胺的恶苗病菌,且均有很高的抗性水平;郑睿等[19]研究发现,江苏省分离得到的水稻恶苗病菌株对咪鲜胺全部达到了中抗和高抗药性水平,其中高抗咪鲜胺的菌株占比已达76.62%,说明咪鲜胺高抗药性菌株已经成为江苏省恶苗病菌的优势群体。
目前,我国水稻恶苗病咪鲜胺抗性研究以江苏省为主,浙江省相关报道不多。本研究自浙江省杭州市富阳区采集水稻恶苗病菌株,对其咪鲜胺敏感性进行检测,以期初步探明当地水稻恶苗病菌对咪鲜胺的抗性水平,为生产上合理用药及相关研究提供参考。
1 材料与方法
1.1 水稻恶苗病病原菌分离
取水稻恶苗病发病茎秆,剪成0.5 cm左右的小段,在0.1%升汞溶液中浸泡消毒1 min,75%乙醇溶液消毒30 s,无菌水漂洗3次后,放在添加硫酸链霉素的水-琼脂培养基(WA)上,每个标本放在一个培养皿里,28 ℃培养至有菌丝产生。挑取菌丝尖端,转移到马铃薯葡萄糖琼脂培养基(PDA)上,培养直至产孢,经单孢分离后,得到水稻恶苗病菌株。
1.2 咪鲜胺对各菌株的毒力测定
参照行业标准NY/T 1156.2—2006《农药室内生物测定试验准则 杀菌剂 第2部分:抑制病原真菌菌丝生长试验 平皿法》测定咪鲜胺对各菌株的毒力,略有改动。在无菌操作条件下,根据试验处理将预先融化的灭菌培养基定量(一般为90 mL)加入无菌锥形瓶中,从低浓度到高浓度依次定量吸取药液,分别加入上述锥形瓶中,充分摇匀。然后等量倒入3个以上直径9 cm的培养皿中,制成相应浓度的含药平板。试验设不含药剂的处理作空白对照,每个处理不少于3个重复。将培养基培养的恶苗病菌(菌龄5 d左右),在无菌条件下用直径5 mm的灭菌打孔器自菌落边缘切取菌饼,用接种器将菌饼接于含药平板中央,菌丝面朝上,盖上皿盖,置28 ℃培养箱中培养。培养7 d后调查病原菌菌丝生长情况。用卡尺测量菌落直径(mm)。每个菌落用十字交叉法垂直测量直径各一次,计算各处理浓度对供试靶标菌的菌丝生长抑制率(%)。
1.3 水稻恶苗病菌的抗药稳定性
在获得咪鲜胺对各菌株半数效应浓度(EC50)的基础上,分别选择3个敏感菌株和4个高抗药性菌株,接种PDA平板,每2~3 d转接一次,称为一代。分别测定咪鲜胺对转接5、10、20代后敏感菌株和高抗药性菌株的EC50。
1.4 水稻恶苗病菌对咪鲜胺和其他杀菌剂的交互抗性
在获得咪鲜胺对各菌株EC50的基础上,分别选择2个敏感菌株和2个高抗药性菌株。分别测定嘧菌酯、多菌灵和戊唑醇对以上4个菌株的EC50,分析咪鲜胺和上述3种杀菌剂的交互抗性。
1.5 数据分析
利用DPS v9.0.5数据分析软件,根据各药剂浓度对数值及对应的菌丝生长抑制率作回归分析,计算各药剂的EC50。
2 结果与分析
2.1 水稻恶苗病菌标本的采集和分离
共采集水稻标本106份,包括水稻品种60种以上,分离得到单孢菌株85株。
2.2 水稻恶苗病菌对咪鲜胺的敏感性
通过菌丝生长抑制法测定85个水稻恶苗病菌株的EC50(图1),值域为0.012~0.620 mg·kg-1,敏感性相差51倍,但多数菌株的EC50在0.05~0.10 mg·kg-1。
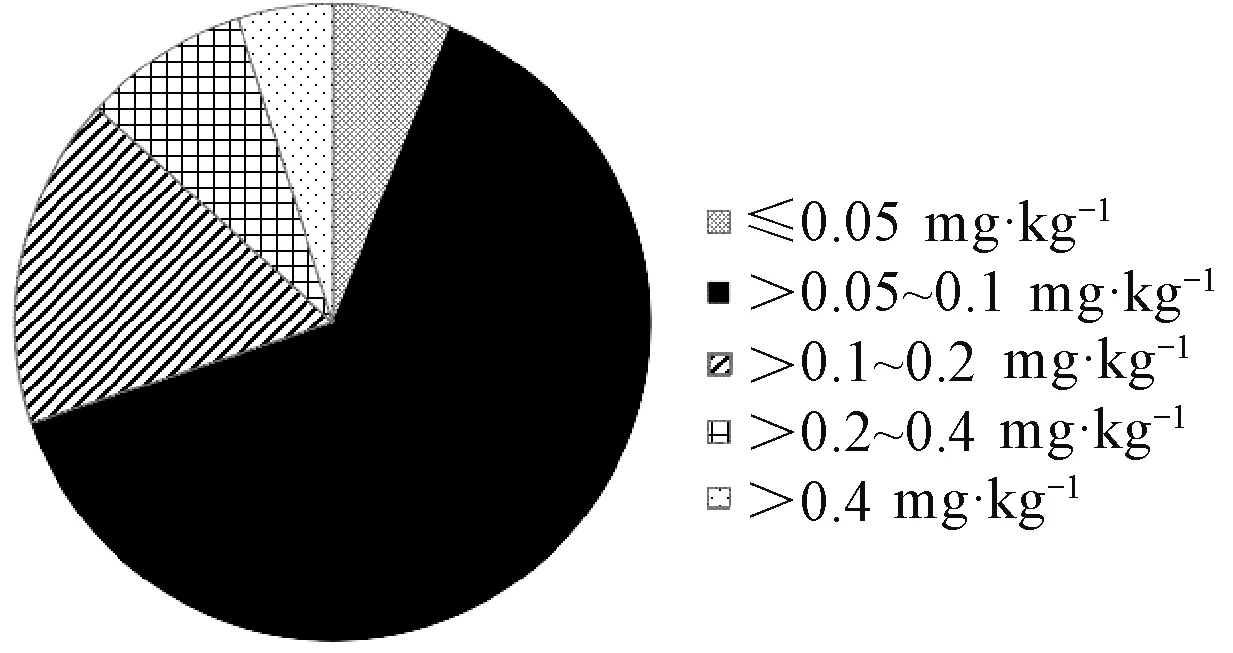
图1 各菌株EC50的分布
2.3 水稻恶苗病菌株的抗药稳定性
选取3个最敏感菌株和4个抗药性最强的菌株,分别转代培养5、10、20次,开展抗药稳定性测试。结果(表1)表明,敏感菌株的EC50随着转接代数升高差异并不明显,说明敏感菌株在无药剂选择压力的情况下,敏感性保持稳定;而高抗药性菌株对咪鲜胺的抗性随着转接代数提高呈明显下降趋势,其中,CR074转接20代后,其EC50从0.530 mg·kg-1下降到0.370 mg·kg-1,其他3个高抗药性菌株转接20代后,EC50仍维持在0.400 mg·kg-1。上述结果表明,水稻恶苗病菌对咪鲜胺的抗药性并不稳定,在无咪鲜胺选择压力的情况下,其抗药性可随着菌株的自然繁殖而逐渐降低,但这需要相当长的时间。
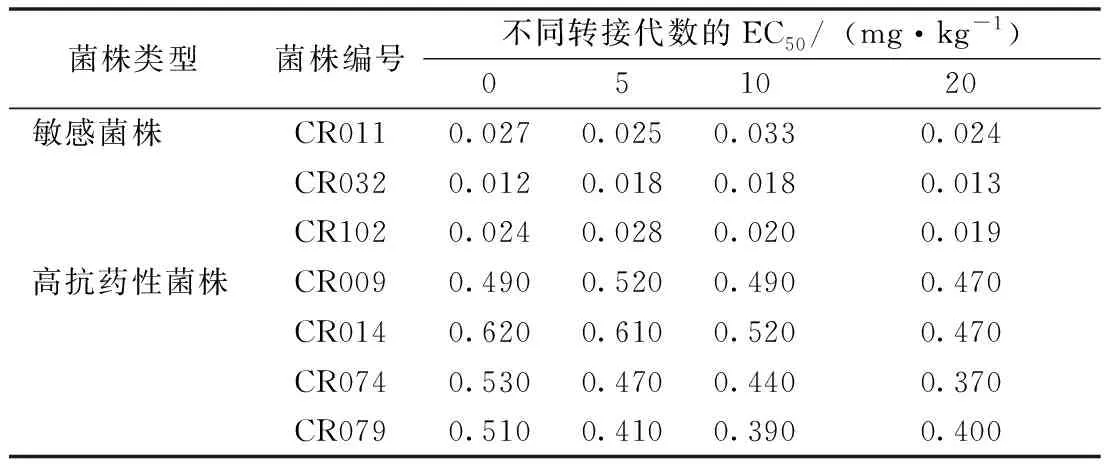
表1 水稻恶苗病菌株对咪鲜胺抗药性的稳定性
2.4 水稻恶苗病菌对咪鲜胺和其他杀菌剂的交互抗性
从表2可以看出,咪鲜胺敏感菌株(CR032和CR102)和高抗药性菌株(CR014和CR074)对嘧菌酯的EC50分别为0.71、0.45、0.52、0.24 mg·kg-1,说明水稻恶苗病菌对咪鲜胺和嘧菌酯无交互抗性。咪鲜胺敏感菌株(CR032和CR102)和高抗药性菌株(CR014和CR074)对多菌灵的EC50分别为1.71、1.45、1.61、1.42 mg·kg-1,说明水稻恶苗病菌对咪鲜胺和多菌灵亦无交互抗性。咪鲜胺敏感菌株(CR032和CR102)和高抗药性菌株(CR014和CR074)对戊唑醇的EC50分别为0.037、0.024、0.080、0.110 mg·kg-1,咪鲜胺高抗药性菌株对嘧菌酯的EC50明显高于咪鲜胺敏感菌株,说明水稻恶苗病菌对咪鲜胺和戊唑醇有交互抗性。
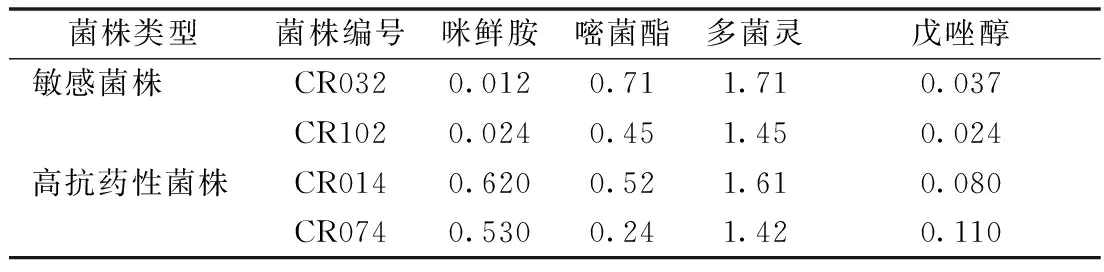
表2 水稻恶苗病菌株对咪鲜胺及其他 杀菌剂的EC50 mg·kg-1
3 讨论
目前,已有多个关于水稻恶苗病菌株对咪鲜胺的敏感性研究,但是研究结果差异很大,可能与培养基、培养温度、培养时间等因素有关。EC50值并不是绝对的,它受很多因素的影响。本研究通过菌丝生长抑制法测定了85个水稻恶苗病菌株的EC50,值域在0.012~0.620 mg·kg-1。相对其他研究,本研究采样范围小,但样本量大,得到单孢菌株85株。在本研究检索范围内,还未见对小范围的水稻恶苗病抗药性分化的研究,本研究的完成可为小范围内水稻恶苗病菌及其他病菌抗药性分化的研究提供参考。
杀菌剂的敏感性在长期停用同一作用类型的杀菌剂后,可以得到一定恢复,其恢复时间和恢复程度与病原菌对该类杀菌剂的抗性稳定性有关[20]。水稻恶苗病菌对咪鲜胺的抗药性并不稳定,在无咪鲜胺选择压力的情况下,其抗药性可随着菌株的自然繁殖而逐渐降低,但这需要相当长的时间。
作物病原菌对药剂的交互抗性是指对一种杀菌剂有抗性的菌株,同时对另外几种杀菌剂有抗性的现象。一般认为同一作用机制的杀菌剂容易产生交互抗性。本研究表明,咪鲜胺和戊唑醇具有交互抗性,而与多菌灵和嘧菌酯无交互抗性。关于上述交互抗性机制,还有待于进一步研究。
猜你喜欢
世界农药(2022年8期)2022-09-02
新农民(2020年5期)2020-12-10
农药科学与管理(2019年7期)2019-11-29
农药科学与管理(2019年7期)2019-11-29
今日农业(2019年14期)2019-09-18
世界农药(2019年3期)2019-09-10
农药科学与管理(2019年5期)2019-08-13
世界农药(2019年2期)2019-07-13
农药科学与管理(2019年10期)2019-04-20
营销界(2015年23期)2015-02-28
