
猪场咬尾行为早期自动监测预警研究(下)
2019-03-11 09:28张振玲编译
猪业科学 2019年2期
张振玲 (编译)
(徐州生物工程职业技术学院动物工程系,江苏 徐州 221006)
2 结果
2.1 咬尾暴发情况
断奶后(16~41 d)(平均f标准误差=28.2f9.3),23组不断尾猪中有15组(占65.2%)暴发咬尾,其中有5起咬尾发生在断奶圈,10起在保育圈。
2.2 咬尾损伤随时间推移的评分情况
共记录12 440个咬尾损伤。图1显示了暴发组平均咬尾损伤评分在咬尾暴发前后数周的占比情况。
咬尾暴发前几周,伤害和新鲜度分数开始增加,而肿胀和尾部长度分数仅在暴发之前或之后才增加。在暴发前后2周,非暴发组猪的损伤评分为0的平均比率仍保持在0.6~0.7(暴发前2周为0.677f0.064;前1周为0.663f0.061;暴发1周后为0.709 9f0.067 1;2周后为0.592 9f0.069 3),这一比例在暴发组显著下降(暴发前2周为0.556f0.061 6,前1周为0.375f0.375;暴发1周后 为 0.198f0.062 7,2周后为 0.206 5f0.068 2)(图1)。线性混合模型揭示了不断尾处理的咬尾损伤极为显著(F1,7= 24.98,P= 0.002)、不断尾处理周数(F3,50= 9.53;P< 0.001)以及两者之间的交互作用均极为显著(F3,50= 8.01,P< 0.001)。LSD 方差分析显示对照组与处理组比较,除暴发前2周差异不显著外,其余周差异均显著(P<0.05)。

图1 15组不同咬尾损损伤评分在咬尾暴发前后数周平均占比图
2.3 3D数据采集
收集到的3D数据为1 200 d(23组h52.2 d),但由于各种技术原因,仅收集962 d,其中827 d用于分析(其中有135 d尾姿数据不到100,故分析中舍弃)。通过机器视觉算法,共获得2 152 101个3D尾姿测量数据结果,但每天的数量变化很大。1 d中数据最多为20 371,平均(±标准误)为2 237(±2 746)。其中,尾角测量值为0˚时的占58.2%(3D 0;1 251 601次),表明尾姿过低,不能检测到角度。角度0˚~30˚占19.3%(3D 1;416 250 次 ),30˚~ 60˚占 14.9%(3D 2;321 552次 ),60˚~ 90˚占7.6%(3D 3 162 698次 )。
2.4 2D视频数据进行3D数据验证
3名观察者共产生926个3D尾姿数据(图2)。该算法擅长识别低靠身体的尾部(并且很好地识别低悬垂尾部),均属低尾姿。人眼识别为卷尾(302头)的被3D算法准确识别为低尾姿267头(3D 0),占88.4%(267/302)。人眼识别为未卷尾(624条)被准确地识别为尾姿不低(3D 1,2或 3),占66.8%(417/624)。卡方检验显示卷尾与3D 0(低尾姿)极显著相关(χ2=248.5,P<0.001)。因此,该算法的总体准确率(正确识别684/总926)为73.9%。对于低尾姿,该算法识别为3D 0的准确率为53.4%(71/133)。若将卷尾和下垂尾的视觉识别合起来,将其识别为3D 0,则该算法的灵敏度为77.7%,特异性为72.3%,准确率为74.8%。在检测到的474次3D 0低尾姿中,有71.3%属卷尾或低姿松弛,高尾姿有56.2%(50/89)被正确识别为3D 2或3D 3。尽管经常观察到卷尾(386/926=41.7%的人眼检查率),但3D算法处理得不太好,在所有3D类别中识别率相当均匀(图2)。
2.5 咬尾暴发群前后3D低尾姿情况

图2 人眼检查2D数据(纵坐标)验证3D尾姿数据(横坐标)的条形图
在15个暴发组中,绘制了相对于咬尾暴发日的3D 0低尾姿日占比(图3)。每组可用数据25 ~ 51 d( 平 均 41.93f7.95 d),拟合的简单多项回归模型具有极显著 意 义(F2,543=101.33,P <0.001;R2=26.9;表2)线性项和二次幂项均有极显著性差异(线性估计为33.2h104,P <0.001;二次幂项估计为 -2.5h104,P <0.001)。正的线性项表示3D 0(低尾姿)随时间推移而增加,而负二次项则表示数据正在下降(呈倒u形;图3),反映了随咬尾暴发临近,低尾姿比例增加,然后在采取各种缓解措施以减少咬尾的情况下,暴发后低尾姿下降。拟合第二类回归模型,每组拟合单条直线(F44,501=42.97,P<0.001),拟合效果较好(R2=77.2)。在15个咬尾暴发组中,有13组线性关系显著(P<0.05),其中9组呈正相关,3 D 0(低尾姿)随时间推移而增加,4例呈负相关。11组具有显著的二次幂(非线性相关)(P<0.05),其中10组具有负的二次幂系数,表明3D 0(低尾姿)与时间呈倒 U 关系(图3)。一组有一个正二次项表示 U 形的。
2.6 暴发组中咬尾暴发前3D 0低尾姿情况
对15个暴发组,每一天低尾姿(3D 0)的比例与暴发前的数据进行拟合(用于分析的数据如图3所示,位于y轴的左侧),在此分析中,每组有1~37 d的可用数据(平均20.87f9.87)。这些数据拟合的简单多项回归模型间具有显着的相关性(F2,243=54.3,P <0.001;R2=30.3)。正线性项(193.3h104,P <0.001)表明低尾姿猪的比例在咬尾暴发前有所增加,而二次正相关系数(3.6h104,P<0.001)则呈上升趋势。模型拟合各组的独立直线,总体拟合度较好(F41,204=32.4,P<0.001;R2=84.0),其中 8条呈显著正线性相关(6条不显著),8条具有显著的正二次系数(非线性,二次函数))。
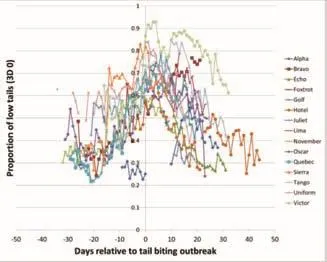
图3 咬尾暴发前后数天不同组的低尾姿(3D 0)的比例
2.7 暴发组中咬尾暴发前1周和2周3D低尾姿比较
有足够资料可用于分析的10个暴发组(第2周和第1周至少有2天的数据),低尾姿猪的比例(3D 0)在暴发前第1周(0.562f0.009)极显著高于暴发前第 2周(0.473f0.009,F1,107=47.5,P <0.001;图 4)。
此结果得到了同一数据回归分析的支持(极显著)(F1,107=67.6,P <0.001)。 正 的 回 归 系 数(0.013f0.002)表明在暴发前14 d低尾的比例增加。
2.8 暴发组与对照组间3D低尾姿的比较
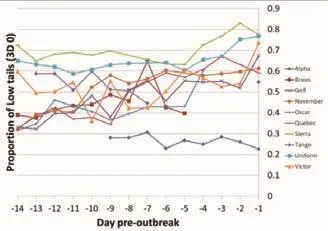
图4 在暴发前的若干天检测到低尾姿(3 D 0)的比例
采用线性混合模型,比较各批次内暴发组和对照组在暴发前第2周、前第1周、暴发后第1周、后第2周的咬尾发生情况。在校正了生长阶段对咬尾暴发极显著影响(断奶阶段=0.53f0.09,保育 阶 段 =0.63f0.09;F1,215=126.5,P <0.001)后,暴发组猪的低尾姿率显著高于对照猪(F1,5=7.47,P =0.046;图5)。最小显著性差异检验显示,暴发组与对照组猪在咬尾暴发前第1周、暴发后第1和第2周间的低尾姿差异显著(P <0.05)。有极显著的时间处理交互作用(F3,93= 29.5,P< 0.001),低尾姿随时间的推移而增加,而对照组则不是。
2.9 3D低尾姿作为损伤评分预测指标
采用线性混合模型,确定各组(包括暴发组和对照组)的3D低尾姿与尾部损伤的总体关系。表2中的负效应系数表明,当低尾姿较少时(3D 0),有较多未受伤尾(损伤0、新鲜0、肿胀0)或轻微损伤尾(损伤1或2)。相比之下,表2中的正的系数表明,较低的尾部(3D 0)预示更大比例的受损尾部(损伤评分3或4)、新鲜受伤的尾部(新鲜度评分1、4或5)或减少的尾部长度(长度1)。
3 讨论
咬尾在某种程度上是不可预测的,何时暴发仍然是变化和难以预测的。

图5 咬尾暴发前2周和后2周3D低尾姿(3D 0) 占比的估计均值(±标准误)
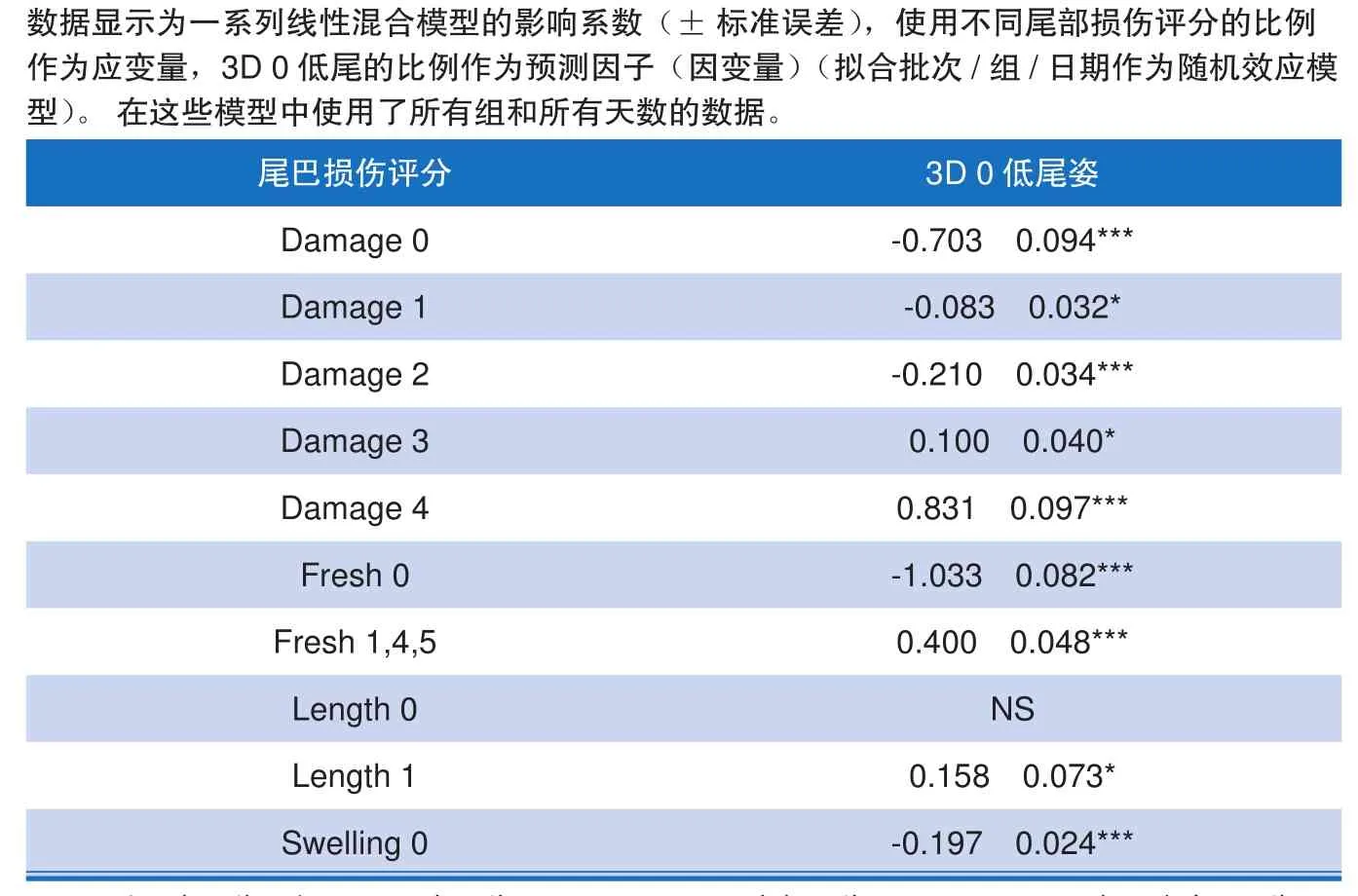
表2 尾部损伤评分与3D低尾数据的关系
本研究第1个目的是用观察者对尾部姿态的视觉评估来验证3D尾部姿态检测算法。算法性能并不完美,而且对于最常见的尾部姿态如尾巴卷曲,显然算法还有改进的余地。只有3D 0是足够可靠的可以用于进一步分析。第2个目的是确定是否自动检测到(3D 0)低尾姿在咬尾暴发前发生变化。回归分析表明,低尾姿率在咬尾暴发前有所增加且呈上升趋势,在暴发后再次下降。当对咬尾暴发前的2周进行比较时,有证据表明,从前2周到前1周,以及在14 d内,低尾姿数都在增加。这些结果也为其他作者所提出的咬尾暴发前低尾姿势确实增加提供支持,并可用作咬尾暴发的预警信号。第3个目的是比较咬尾暴发组和非暴发组(对照组)的3D尾部姿势数据。在咬尾暴发前前1周和暴发1周和2周后,即使考虑了不同生长阶段的影响,暴发组的低尾率也高于对照组。这为使用这种技术作为咬尾预警信号的想法提供了进一步的支持。尽管对照并非完全没有尾巴受伤,但这些暴发组与对照组的低尾姿确实存在差异。增加暴发组和对照组之间尾部损伤程度对比的方法是使用高水平的环境富集材料,以降低对照组咬尾风险。然而,这种环境变化本身可能会影响到尾巴姿势以致于引起困扰,而且咬尾的不可预测性和群体间的变异性可能仍然会导致一些尾巴损伤和环境富集的对照圈咬尾的暴发。第4个目的是确定不同时间和不同组群的3D尾部姿势数据与尾部损伤评分之间是否存在普遍的关系。这一分析忽略了暴发组和对照组之间的区别。一系列线性混合模型显示3D尾部姿势数据与尾部损伤评分之间有显著的关系:当组中有许多未受伤的猪,或轻度受伤的猪时,检测到有较少的低尾姿(3D 0)。当该组中有更多受伤的猪时,低尾姿比例更大。图3中的倒“U”图也反映了随着暴发越来越近而低尾姿增加,以及当采取措施停止进一步的咬尾以使尾巴(损伤)恢复时,咬尾暴发后低尾姿下降。
有些研究发现在咬尾暴发之前,猪的活动增加了。因此,猪的活动情况也有作为预警信号的潜力。从原理上讲,每天3D尾巴检测的数量可以作为猪行为活动的代表,因为猪必须站在摄像机下才能被检测到,但是由于每天检测到数据变化很大,也无法分析这一点。检测到尾姿次数的多变性有各种可能的原因。所有的摄像头都连接到一台PC上,因此系统可同时从所有圈中获取数据。相机所能捕捉到圈里猪的数量可能会有所不同。猪尾部运动量(如尾部摆动)可能在几天内有所变化在,这可能会影响到检测数据。在整个项目进行过程中,对数据捕获和尾部姿态检测算法作了一些优化,因此不可用而舍弃的数据也减少了,并在以后的批次中检测到的数据量也增加了,但每天的检测次数仍然是可变的。
本项研究就是找出猪早期咬尾信号,这个信号对于猪农来说要足够早以便识别咬尾暴发迹象并采取措施。本研究咬尾暴发的标准是:在圈外确认有3头有新鲜伤口的猪,或者一头或多头尾巴流血,或者有明显的咬尾行为。尾部评分数据(基于从圈内对尾部的密切观察)显示,至少暴发前1~2周,尾部损伤的迹象逐渐增加(图1)。其他研究者对暴发使用了不同的定义或标准。Zonderland等人分析了单头猪只水平的数据,并表明猪尾巴在从没有损坏到小孔咬痕的过渡过程中,以及在从未受伤或有咬痕的尾巴过渡到有明显的尾部损伤时,更有可能夹在两腿之间。Lahrmann等采用的标准是每圈内出现4头有受伤的猪(包括正在恢复的及擦伤)。若我们使用Lahrmann等的定义,很可能会比我们的方法更早地在某些圈暴发咬尾。 早期 信号中早期的概念显然与 咬尾 的定义有关。在咬尾暴发之前就有明显的尾巴伤害(因为在尾巴咬伤暴发之前就有仔细的尾巴打分),这表明如果结合尾部姿势能够开发一种自动检测尾部损伤的方法(如尾巴颜色或尾部温度),也可以提供咬尾暴发的早期预警。
各组(图3、图4)之间的低尾姿(3D 0)占比差异较大,这也反映在改进的每组拟合回归模型中。从3D数据预测咬尾暴发可能是监测咬尾的较好方法。有时,随着时间的推移,尾巴姿势的趋势在同龄群体之间会发生变化,可能与咬尾无关。特别是,当低尾姿(3D 0)的猪被转到新圈时,它们在对照组和暴发组中的比例往往会同时显著增加。显然,我们还需要了解影响这一 本底 尾姿的因素,这将有助于进一步完善预警系统。例如,尾巴姿势可能是猪的唤醒状态时或情绪状态的普通信号,这意味着它可能被其他生理或社会压力源或疾病状态所改变,而不仅仅是早期咬尾的信号。另一担忧是,咬尾风险的多因素性最有可能意味着某些类型的暴发可能比其他类型更容易被早期发现。如果由饮水、料槽、通风或供暖故障引起的应激而导致的咬尾暴发,可能会相对更快地发生。
本研究中尾姿变化和尾部损伤暴发前的变化与其他研究者所发现的类似。这些变化加上尾部受伤情况(即使在缺乏高科技手段的情况下),也可以作为猪农的咬尾暴发早期预警信号(在有足够时间密切和频繁地检查他们的猪的情况下)。提高对早期预警信号的认识和使用可以减少咬尾的不可预测性,使养猪者更有信心遵守欧盟理事会第2008/120/EC号指令的要求,停止使用断尾作为预防咬尾的 杀手锏”。
本研究的发现为以下设想提供了 理论证据”:使用3D摄像机来记录尾部姿势可以发展成一种预警系统,以防止咬尾暴发。这一技术能否商品化,将取决于实时预测系统能否成功设计,并取决于该系统的经济成本和效益。3D相机的成本相对较高,但如果除咬尾预测外还用于检测猪的生长、攻击行为或健康不良行为指标等其他重要经济特性等多功能系统,则使用3D相机在经济上更为可行
本研究首次表明,利用Time-of- fl ight 3D摄像机和机器视觉算法检测低尾姿有望提供猪咬尾自动预警信号。尾姿检测算法具有足够的准确性,低尾姿率随时间的推移而增加,在暴发组中高于对照组,且与尾部损伤增加有关。本研究通过提供有关动物生长、健康、行为和福利的实时信息,为快速发展的 精确畜牧养殖业 领域做出贡献,利用新技术为猪场管理决策提供信息。(全文完)
猜你喜欢
舰船科学技术(2022年20期)2022-11-28
文苑(2020年5期)2020-06-16
小学生学习指导(低年级)(2020年3期)2020-06-02
红领巾·探索(2019年6期)2019-08-01
学生导报·东方少年(2019年28期)2019-01-17
中国生殖健康(2019年10期)2019-01-07
学苑创造·A版(2017年8期)2017-06-24
作文周刊·小学一年级版(2017年9期)2017-06-20
小学阅读指南·低年级版(2016年7期)2016-05-14
