
常春二乔玉兰春夏季开花节律及营养效应研究
2019-03-19 08:50孙李勇刘晨妮尹增芳
植物研究 2019年2期
蒋 政 孙李勇 刘 旭 刘晨妮 尹增芳,2*
(1.南京林业大学南方现代林业协同创新中心,南京 210037; 2.南京林业大学生物与环境学院,南京 210037)
在自然界中,生长在温带的植物通常一年只开一次花,但是也有一些植物存在多次开花的现象。一般认为果木类一年多次开花会导致减产现象的发生[1],而园艺植物多次开花则可以增加观赏性状的多选择性。王俊杰和汪琪华[2]最早总结了生长在我国的木本植物二次开花的类别,但关于植物二次开花现象机理的解读资料较少。在黄牡丹杂种(Paeonia‘High Noon’)二次开花现象研究中,发现其生长发育年周期、花芽分化[3]及二次成花的酶代谢具有一定规律性[4]。发育生物学已经证实,碳水化合物、蛋白质等营养物质的分配格局与开花密切相关。郭娜[5]认为3种丁香属(Syringa)植物二次花芽分化阶段营养物质发生规律性变化,而在砂梨(Pyruspyrifolia)第二次开花过程中,枝条内有机营养物质含量的变化亦非常显著[6]。因此,比较不同开花期营养物质的动态变化状况,更有助于对二次开花现象的理解。
常春二乔玉兰(Magnoliasoulangeana‘Changchun’)属于木兰科(Magnoliaceae)木兰属(Magnolia),是由二乔玉兰(Magnoliasoulangeana)芽变枝繁育而来,具有与二乔玉兰同样的观赏性状,如早春开花,先花后叶,花繁茂艳丽等,但相对于二乔玉兰,在同样的栽培环境下,常春二乔玉兰一般一年至少开花两次[7],而二乔玉兰则仅于春季开花一次。在自然生长条件下,常春二乔玉兰与二乔玉兰一样经历营养生长向生殖生长阶段的转变,但成花后的常春二乔玉兰可分别于春、夏季各开花一次。这一现象值得人们进一步的思考:常春二乔玉兰春季开花与夏季开花特性有何差异?是怎样的营养水平及其效应机制可以导致其两季开花?基于上述问题,本研究拟对常春二乔玉兰春夏两季开花现象进行形态及营养物质含量变化的观测及分析,为木本植物多次开花的生物学研究提供基础理论资料。
1 材料与方法
1.1 试验材料
以南京林业大学校园中6年生的常春二乔玉兰为试验材料,树体无病虫害,常规养护管理。
1.2 试验方法
1.2.1 年生长物候期划分
自2015~2017年共进行了3次年生长物候期观测。采用常规形态检测的方法,选取生长状态良好的90个枝条进行跟踪标记,测量并记录标记枝上芽的长、宽和生长状态。同时,随机选取树冠中部5枚芽在Nikon SMZ18体视显微镜下进行解剖并拍照,根据标记枝上芽的生长量和发育状态,确定常春二乔玉兰的生长发育物候期,并于开花期统计测算开花率:
开花率(%)=开放花朵数量/花芽总量×100%
(1)
1.2.2 生理指标测定
在明确常春二乔玉兰年生长物候期的基础上,自2017年1月至2017年6月于常春二乔玉兰春夏季开花的各关键物候期进行采样,每次随机采集3~5个生长良好、无病虫害的枝条,清洗擦干,将顶芽下部第2~3节间茎段剪碎混匀作为茎段待测样品。采用蒽酮比色法测定可溶性糖含量[8];酸解法和蒽酮法测定淀粉含量[9];考马斯亮蓝G-250法测定可溶性蛋白含量[8]。试验3次重复。
1.3 数据分析
利用Photoshop CS5软件进行图版制作,物候观测及营养物质测定数据使用Excel 2010、SPSS 21.0进行数据分析与图表绘制。
2 结果与分析
2.1 年生长物候动态
2.1.1 芽发育动态
自1月上旬至2月上旬,常春二乔玉兰标记枝顶花芽长宽无明显变化。2月下旬,花芽发育膨大,其长宽生长量分别高于2月上旬36.29%和24.21%。3月初,花芽生长量进一步增加(图1a),并开放成花。3月下旬,常春二乔玉兰标记开放成花的枝上侧芽开始萌动,并抽出新生枝条,4月至5月下旬新枝顶芽不断生长。对新枝顶芽解剖发现,常春二乔玉兰于5月初完成营养顶端向生殖顶端的转化(图2a),5月中上旬开始分化形成花被片(图2b),5月中下旬开始分化形成雌、雄蕊(图2c),及至5月底花芽形态分化基本完成(图2d)。随后枝顶花芽生长量出现两种不同的变化趋势:一部分花芽发育膨大,其长宽生长量分别高于5月下旬的60.15%和44.44%,并于6~7月渐次开放成花;另一部分花芽则未发育膨大,生长量亦明显小于膨大花芽。自7~8月,未膨大花芽生长量仅小幅增长。9~12月由于未膨大花芽芽鳞片的脱落,致使其长度有所下降,然其宽度却无明显变化,说明此时未膨大花芽逐渐进入休眠状态(图1b)。
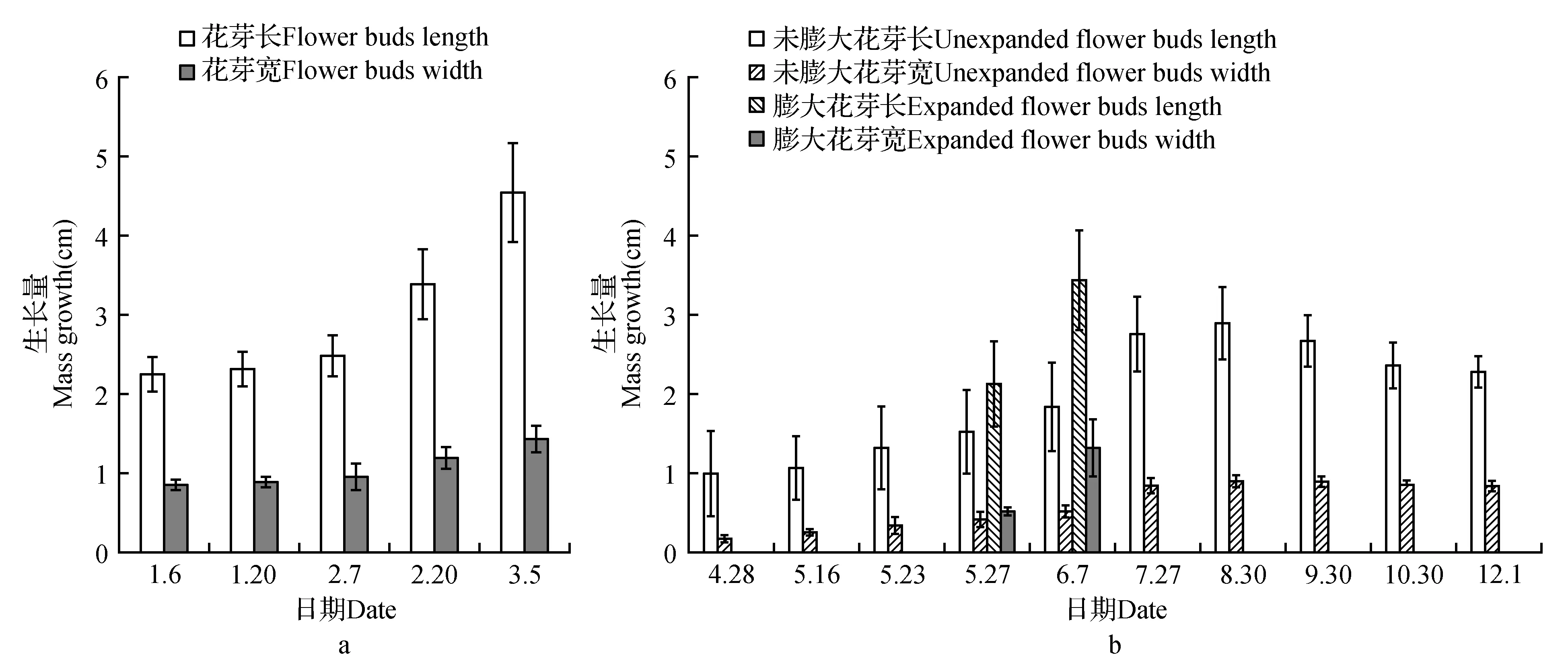
图1 常春二乔玉兰年生长周期顶芽形态变化Fig.1 Morphological changes of top buds during annual growth cycle in M.soulangeana ‘Changchun’
2.1.2 物候期动态
如图3所示,自12月始至翌年2月下旬,以枝顶花芽无明显生长量变化的阶段为常春二乔玉兰休眠期。3月初,花芽芽鳞片破裂,露出红色花被片,常春二乔玉兰进入春季开花期,整个春季开花过程历时约20 d。4~5月,开放花枝上侧芽抽枝生长,为营养生长期。常春二乔玉兰具有顶芽成花的自然属性,在营养生长期后枝条顶芽开始生殖转化。花芽形态分化始于5月初,终于5月底,为花芽分化期。随后花芽的发育进程分为两种类型:迅速膨大花芽与未膨大花芽。6月后,一部分花芽迅速膨大开放成花,常春二乔玉兰进入夏季开花期,历时约20 d。6~7月内未膨大花芽正常发育。7月后未膨大花芽发育较为迟缓,将此时期定义为发育滞缓期。此外,少量夏季开放的花的基部侧芽再次分化形成花芽。10~11月常春二乔玉兰开始落叶,标志着落叶期的到来,枝条叶片全部脱落后,树体进入休眠期。
2.2 春夏季开花模式差异
常春二乔玉兰营养生长后分化的花芽具有花开两季的特点,而且春夏季花期内开花模式存在一定差异。春季开花为先叶开花,而夏季开花则为花叶同放。春季开花表现为树冠上所有花芽全部开放,开花率可达100%,且开花同步率较高,而夏季开花为部分花芽渐次开放,开花率约为30%,且开花同步率较低。春季开花时,花朵基部的侧芽明显膨大(图4a),而夏季开花时,花朵基部以及花枝上的侧芽均相对较小(图4b)。春季开花雌雄蕊发育健全(图4c),为可育花,而夏季开花雌雄蕊干瘪细小,发育不完全(图4d),为不育花。两季开花在花色和花形上并无明显差别(图4:a~b)。

图2 常春二乔玉兰花芽分化过程 a.生殖转化初始阶段;b.花被片分化阶段;c.雌雄蕊分化阶段;d.形态分化完成阶段 RC.生殖顶端;Rec.花托;TeP.花被片原基;SP.雄蕊原基;PP.雌蕊原基;Sta.雄蕊;Pis.雌蕊Fig.2 The process of flower buds differentiation of M.soulangeana‘Changchun’ a. Initial of reproductive transformation; b. Tepal differentiation stage; c. Pistil and stamen differentiation stage; d. Completion of flower bud differentiation RC. Reproductive cone; Rec. Receptacle; TeP. Tepals primordium; SP. Stamen primordium; PP. Pistil primordium; Sta. Stamen; Pis. Pistil

图3 常春二乔玉兰年物候期Fig.3 The annual phenological period of M.soulangeana ‘Changchun’
2.3 春、夏季开花期营养物质变化动态
2.3.1 春季开花期营养物质动态
自休眠期至春季开花期,着生常春二乔玉兰花芽的枝条中,可溶性糖、淀粉和可溶性蛋白含量均总体呈下降趋势(图5)。在花芽开始膨大时,可溶性糖含量下降最为明显(P<0.05),而可溶性蛋白含量则仅有轻微下降,淀粉含量变化不显著,说明可溶性糖的大量消耗为花芽的膨大生长供应了必须的物质储备。随着春季开花期的到来,可溶性糖、可溶性蛋白含量明显下降, 至花期结束时二者降至最低,分别相对于休眠期下降了56.94%和40.34%,淀粉含量同样下降显著(P<0.05),推测春季开花及随后的侧芽萌动需要消耗大量的营养物质。

图4 春夏季开花期间花部形态差异 a.春季开放的花及其侧芽形态;b.夏季开放的花及其侧芽形态;c.春季开放花的雌雄蕊形态;d.夏季开放花的雌雄蕊形态Fig.4 Flower morphological differences during spring and summer flowering periods a. Morphology of flowers bloomed in spring and their lateral bud; b. Morphology of flowers bloomed in summer and their lateral bud; c. Pistil and stamen morphology of flowers bloomed in spring; d. Pistil and stamen morphology of flowers bloomed in summer

图5 春季开花期营养物质含量变化Fig.5 The nutrient contents during spring flowering

图6 夏季开花期营养物质含量变化Fig.6 The nutrient contents during summer flowering
2.3.2 夏季开花期营养物质动态
自花芽分化期至夏季开花期,可溶性糖和淀粉含量总体呈现“降—升”趋势,而可溶性蛋白则表现为下降趋势(图6)。可溶性糖和淀粉于花芽膨大生长时达到最低值,并分别相对于花芽分化期下降了35.27%和35.12%,可溶性蛋白含量亦明显下降,而此时正值花器官进一步发育与花芽迅速膨大生长阶段,可见夏季开花的启动需要大量营养物质的供给。随着夏季开花期的到来,可溶性糖和淀粉含量均有所上升,而可溶性蛋白含量则继续下降至最低,且相对于花芽分化期下降了33.91%,推测碳水化合物含量的升高与含氮化合物含量的降低有利于夏季开花的实现。
3 讨论
3.1 常春二乔玉兰春夏季开花模式的特异性
常春二乔玉兰营养生长后分化的花芽能够花开两季,这与什锦丁香(Syringa×chinensis)、小叶丁香(Sytingamicrophylla)、蓝丁香(Syringameyeri)[5]的二次开花特性类似,而与黄牡丹杂种[3]的一年多次花芽分化而多次开花的特点略有不同。另外,有些植物如苹果(Maluspumila)[1]、梨[10]、桑树(Morusalba)[11]等会因为人工处理、气候变化、病虫害或养护不当而导致二次开花,这些二次开花现象往往不稳定,甚至因营养供应不均而有损经济效益,而常春二乔玉兰的春夏季开花乃是品种遗传特性使然,开花物候期稳定。相较春季开花的白玉兰(Magnoliadenudata)和二乔玉兰,它具有夏季花叶同放的特性,而相较于夏季开花的广玉兰(Magnoliagrandiflora),它又具有春季先叶开花的特性,故此常春二乔玉兰因其独特的物候特性和开花优势而具有更高的观赏价值。
从开花育性角度而言,荚蒾属(Viburnum)、绣球属(Hydrangea)以及菊科(Asteraceae)部分物种[12]的可育花与不育花出现于同一花期,且二者分布于花序的不同位置,属于空间上的育性差异,而常春二乔玉兰的可育花出现于春季开花期,不育花则出现于夏季开花期,属于时间上的育性差异。夏季开花不育虽不具备明显的生殖生物学意义,但却在降低了开花消耗的同时,仍保证了一定的观赏性,大大增加了常春二乔玉兰的观赏价值。
3.2 春、夏季开花模式与营养物质的动态平衡
3.2.1 先叶开花模式与营养物质的再分配
花芽在不同发育阶段,均是生长代谢中心,要从植物体的不同部位大量调集贮藏的无机和有机营养物质,为花芽生长发育提供碳骨架和能源[13~14]。有研究认为,开花期植物的茎不像其他器官那样能够消耗或积累可溶性糖,而仅有分配和转运营养物质的作用[15],这与本研究相关结果不一致。休眠期常春二乔玉兰枝条的可溶性糖、淀粉和可溶性蛋白含量均较高,说明彼时茎为营养物质的储存器官。春季开花期间,萌动开放的花从枝条中调集所需营养,故枝条可溶性糖和可溶性蛋白含量明显下降。枝条中淀粉含量仅于开花后期下降明显,说明常春二乔玉兰春季开花早期以可溶性糖和可溶性蛋白为主要能量和营养来源,直至春季开花后期,为供应开花和侧芽萌动,致使枝条中储存的淀粉分解。由于先叶开花树种在春季开花时无叶片提供同化产物,故常春二乔玉兰与毛泡桐(Paulowniatomentosa)[16]类似,二者在开花过程中所消耗的营养物质皆来源于树体上一年的营养贮存。
3.2.2 花叶同放模式与营养物质的再分配
花芽膨大生长时,枝顶花芽大量消耗营养物质,进一步分化发育并膨大,成为生长代谢中心,故枝条中3种营养物质均明显下降。由于常春二乔玉兰夏季开花时花叶同放,叶片中的光合产物又具有就近分配的特性[13],所合成的碳水化合物通过枝条向花转运,导致茎中可溶性糖与淀粉含量在开花后均显著上升,这与马铃薯(Solanumtuberosum)[17]开花期的碳水化合物变化趋势基本一致。同期可溶性蛋白含量明显下降,则与‘丰水’梨(Pyruspyrifolia‘Hosui’)[6]第二次开花期蛋白质含量的变化类似,上述结果可能与碳水化合物的合成加速以及蛋白质水解酶活性增强有关。谢寅峰等[18]研究表明鹅毛竹(Shibataeachinensis)开花期间可溶性糖和可溶性蛋白的比值有所升高,且均高于不开花植株,这说明碳水化合物含量的相对升高以及含氮化合物含量的相对降低,有利于植物二次开花的实现。
此外,研究表明,糖可能以某种信号分子的形式存在,以类似植物激素的方式,调控植物生长发育和成熟[19~21],如蔗糖能够促进晚花生态型拟南芥(Arabidopsisthaliana)、油菜(Brassicacampestris)、白芥(Sinapisalba)和葡萄(Vitisvinifera)开花,而蔗糖介导的信号途径可能分别在CONSTANS下游和FLOWERINGLOCUST上游,通过整合到光周期途径中发挥作用[22];海藻糖-6-磷酸亦对生长发育有重要作用[23],其合成酶缺失突变体能显著延迟拟南芥开花[24];拟南芥淀粉缺失、合成或降解突变体的开花时间亦会被显著推迟[25~26]。因此,夏季开花期可溶性糖和淀粉含量的提高是否还作为某种信号分子,参与常春二乔玉兰夏季开花的调控则有待于进一步研究证实。
综上所述,在常春二乔玉兰春夏季开花期内,营养物质的分配与消耗因开花模式不同而相异。枝条不仅是营养物质的运输器官,也兼具储藏功能。春季开花以营养物质的消耗和利用为主要模式,夏季开花则不仅仅需要营养物质的充分供给,其动态平衡也对此时开花的实现具有重要的作用。
猜你喜欢
中国农学通报(2022年29期)2022-11-25
中国果业信息(2022年4期)2022-11-22
茶道(2022年3期)2022-04-27
国外畜牧学·猪与禽(2022年1期)2022-04-23
今日农业(2021年7期)2021-07-28
今日农业(2021年3期)2021-03-19
现代农业科技(2018年22期)2018-01-15
中学生理科应试(2017年7期)2017-08-09
绿色科技(2017年7期)2017-05-12
江苏农业科学(2016年12期)2017-04-05
