
环境污染物对精子损伤机制的研究进展
2019-04-17 06:15荆晶于淼汝少国
生态毒理学报 2019年6期
荆晶,于淼,汝少国
中国海洋大学海洋生命学院,青岛 266003
生殖系统的健康是生物体种群得以延续和繁衍的必要条件。然而,日益严重的环境污染问题已经成为包括人类在内的多种生物生殖力下降的元凶之一[1]。精子是雄性生物生殖细胞系发育的终端产物,但在其形过程中,基本丧失了细胞自我修复的机能,因此,更容易受到环境因素的胁迫[2-3]。有报道称,中国男性正面临“精子危机”。在近15年间,中国男性精液质量呈下降趋势[4],且从1995—2008年间精子数量平均每年以2.15%的速度降低[5]。环境污染物不仅会引起精子形态结构的变化如精子畸形、质膜破裂等,还会使精子的数量大幅度减少,造成雄性弱精、少精和无精[6-7]。与此同时,很多污染物还可能通过“攻击”DNA(如导致DNA片段化[7]、形成DNA加成物[8]、改变DNA的表观遗传学修饰[9]等)而改变精子所携带的遗传信息。笔者将从环境污染物诱导氧化应激、改变生精细胞的细胞周期和诱导正常的细胞发生细胞凋亡以及改变生物体雄激素水平等方面综述其损害精子质量和降低精子数量的潜在作用途径。
1 诱导氧化应激(Induce oxidative stress)
动物精子的形态一般可分为3个部分:头部、颈中部和尾部,头部主要由浓缩的细胞核和顶体组成,核内含遗传物质DNA,顶体内则含多种水解酶;颈部连接了精子的头和尾部,由中心粒衍生而来;尾部又可分为中段、主段和末段,中段外包线粒体鞘,主要作用是提供能量,尾部鞭毛的摆动是精子向前运动所依靠的主要方式[10-11]。在生物繁衍时,正常、完整的形态结构直接影响了精子的受精能力,也决定了生育力的强弱。
当生物体暴露于污染物中时,往往伴随着大量活性氧自由基(ROS)的产生。如一些多环芳香烃和卤代芳香烃(多氯联苯、二噁英等),能够激活包括CYP1A和CYP1B在内的I相代谢酶,迅速地对上述物质进行氧化、水解等反应,以达到解毒的目的。但当污染物浓度过高时,一些副反应的发生可导致电子转移和氧化还原过程解偶联,从而释放出ROS。又如,一些微量金属通过自身化学形态的变化及与氧化还原酶中的半胱氨酸残基的反应而改变酶活性,进而导致ROS的形成[12]。通常在生物体中,ROS的产生和清除之间存在着动态平衡,当机体处于氧化应激状态时,会激活细胞中抗氧化小分子的合成和抗氧化酶的表达来清除细胞内的ROS;而当ROS过量产生时,细胞中的氧化防御系统不足以将其完全清除,就会造成氧化损伤[13-14]。精子作为一种高度分化的细胞,大部分细胞器的丢失使其缺乏有效的生物转化和抗氧化防御能力,加之其含有高浓度的不饱和脂肪酸,极易受氧化胁迫的影响[2]。故污染物会通过诱导机体产生氧化应激,从而损伤精子。如亚砷酸盐能够通过增加大鼠睾丸中ROS的含量,造成脂质过氧化产物丙二醛和羰基化蛋白质发生累积,进而降低了精子活力[15]。过量的ROS可以通过多种途径降低精子质量,主要体现在破坏精子结构及损害精子功能2个方面,如图1所示。
1.1 ROS破坏精子结构
精子的质膜中富含不饱和脂肪酸,包含较多的双键,是ROS最易攻击的靶点之一。ROS可进攻精子质膜中的脂肪酸侧链,夺取碳上的氢原子从而启动脂质过氧化反应,被夺去氢原子的脂质过氧化物又会从相邻脂肪酸侧链中夺取氢原子而加剧脂质的过氧化[16]。大量双键的丢失及脂质过氧化产物的形成,会导致精子膜流动性减弱,甚至造成质膜破损[17]。如铅化合物的暴露能诱导男性精子产生ROS,引发氧化应激,导致精子膜脂质过氧化[18]。壬基酚、心得安和己烯雌酚(DES)共同作用时鲟鱼精子发生氧化应激,从而大幅增加了其精子蛋白质、脂质过氧化水平,破坏了精子膜的完整性[19]。
1.2 ROS损害精子功能
1.2.1 影响精子活力
线粒体是精子产能的主要场所,为精子运动提供三磷酸腺苷(ATP),精子的活力直接受线粒体功能的调控[20]。线粒体的内膜上分布有ATP合酶以及电子传递复合物(Ⅰ、Ⅱ、Ⅲ、和Ⅳ),是真核生物电子传递和氧化磷酸化的关键场所[21]。由于线粒体本身富含多种酶、结构蛋白、膜脂质及核酸等,也极易受到ROS的攻击[22]。ROS可以使线粒体膜发生脂质过氧化,影响膜上电子传递链的功能,进而破坏氧化磷酸化过程,影响ATP的合成[23]。如PM2.5就是通过增加细胞内ROS的含量进而损害线粒体的结构,导致ATP的水平降低,使得能量代谢发生阻塞,导致精子活力下降[24]。
线粒体在进行呼吸氧化的过程中,由于呼吸链中的质子和电子的转移,会在内膜两侧造成质子及其他离子浓度的不对称分布,从而形成线粒体膜电位,该电位是产生ATP的先决条件[25]。因此,当精子线粒体膜电位发生改变时,势必会影响供能,进而影响精子的活力。而ROS一方面可以通过诱导线粒体内钙离子的释放、增加内膜通透性降低线粒体膜电位[26];另一方面,ROS和过氧化脂质的含量升高时也会使线粒体膜电位发生损耗[27]。研究发现,多氯联苯1254暴露人类精子后,精子中ROS的含量显著升高,进而扰乱了钙离子的平衡,导致高线粒体膜电位的精子占比呈剂量依赖性下降,精子活力降低[28]。另有研究表明,亚砷酸盐可通过提高大鼠睾丸中ROS的水平,降低大鼠精子线粒体膜电位,进而影响了电子传递链的功能,最终导致精子活力显著降低[15]。
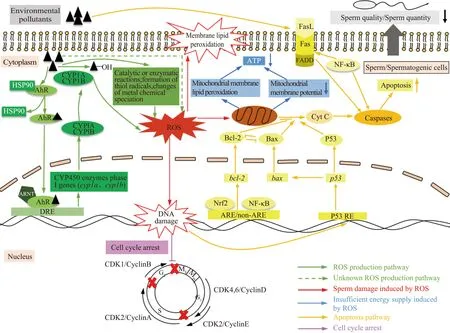
图1 环境污染物通过诱导氧化应激、细胞凋亡以及改变细胞周期损伤精子的机制Fig. 1 The mechanisms of sperm damage induced by environmental pollutants via increasing oxidative stress and apoptosis as well as changing sperm cell cycle
1.2.2 改变遗传物质
除了作用于生物膜,ROS还能够直接攻击DNA中的单个碱基(如鸟嘌呤)或脱氧鸟苷上的C-8位使其氧化,从而造成DNA损伤[29]。此外,ROS所生成的脂质过氧化物也可以共价结合到DNA上,使DNA双链发生断裂,导致遗传信息发生改变或缺失,如铅能通过诱导ROS的产生使精子DNA链发生断裂,造成DNA片段化,导致精子DNA损伤[18]。另有研究表明,相较没有吸烟习惯的男性,长期吸烟的男性精液中ROS的含量显著增加,其精子DNA碎片化指数也随之明显升高[30]。
研究发现,在精子发生过程中,精子DNA损伤与ROS的含量呈正相关关系,而ROS所造成的染色质损伤与精子形态异常之间又高度相关[31]。若在精子头部遗传物质的包装过程中出现DNA突变、DNA损伤和染色体畸变,精子就会表现出异常的形态,导致精子畸形[32]。通常,DNA受损的精子的受精率较低,并且受精后还存在其他问题,如孵化率低、发育异常等[7]。一项野外调查研究中发现,受人类活动污染较重的河水中的软口鱼与轻度污染河水中的鱼相比,DNA受损精子所占的百分比明显增加,同时测得受精卵卵黄囊吸收末期的畸形率显著上升,而子代的孵化率则大幅度下降[2]。
2 改变细胞周期和诱导细胞凋亡(Change cell cycle and induce apoptosis)
据报道,自20世纪90年代初以来,男性精子数量下降一直是全球关注的焦点,环境污染被认为是主要的原因之一[33]。大量的研究表明,污染物可以通过阻碍生精细胞分裂和诱导正常的生精细胞发生凋亡2个方面(如图1所示)降低精子的数量和质量。
2.1 阻滞生精细胞分裂
雄性生物的精子发生过程主要分为3个阶段:精原细胞进行有丝分裂形成初级精母细胞;初级精母细胞经减数第1次分裂形成次级精母细胞,次级精母细胞再经减数第2次分裂形成单倍体精细胞;精细胞经一系列复杂的形态变化发育为具有鞭毛的精子[34]。减数分裂的细胞周期可划分为G0期、G1期、S期、G2期以及减Ⅰ期、减Ⅱ期[35]。无论是初级精母细胞还是次级精母细胞都需要通过减数分裂发育至它们的下一阶段,这个过程也是环境污染物阻碍精子生成的重要靶点。污染物会通过改变生精细胞的细胞周期来阻滞精子的产生。研究发现,PM2.5会损伤小鼠精母细胞DNA,导致细胞周期停滞在G0/G1期,降低了G2/M期的精母细胞数,阻滞了细胞的分裂,最终导致精子数目降低[24]。另有报道表明,香烟烟雾冷凝物可以通过阻断小鼠精母细胞DNA的合成过程而抑制DNA复制,同时下调G1/S期细胞周期蛋白A和E的表达使细胞周期停滞在S期[36]。
还有些污染物通过中断减数分裂的减Ⅰ期进程阻滞精子的产生。如双酚A(BPA)和雌二醇的暴露均能中断大鼠睾丸中细胞减Ⅰ期过程,将减数分裂过程阻滞在了初级精母细胞阶段,显著降低了减Ⅰ期中细线期和偶线期阶段精母细胞百分比,进而减少了精子数量,抑制了精子释放[37]。在小鼠中也观察到了类似的实验结果,由于BPA抑制了精子发生的关键调节因子BOULE的表达,并上调了雌激素受体ERα/β的表达,进而扰乱了减Ⅰ期的进程[38]。此外,dmc1是减Ⅰ期前期表达的特异性基因,其编码的蛋白质参与减数分裂中同源染色的联会[39]。当小鼠dmc1基因被敲除后,其精子发生过程就会停止在减Ⅰ期前期[40]。还有研究表明,DES及氟他胺能显著降低dmc1的表达,从而抑制斑马鱼减数分裂过程,阻碍精子的产生[41]。
2.2 诱导生精细胞凋亡
细胞凋亡又叫作程序性细胞死亡,是细胞处于生理及某些病理状态下,在相关基因的调控下主动程序性死亡的过程[42-43]。正常情况下,细胞凋亡会在精子发生时通过维持生精细胞和支持细胞在数量上的平衡来调节精子的生成[44]。若凋亡异常,必将影响精子的数目及质量进而造成生殖异常。环境污染物则会促进生精细胞发生凋亡,由此影响生精过程,进而降低精子数量。如香烟烟雾暴露会导致男性精子膜内侧磷脂酰丝氨酸翻转至膜外侧,导致DNA双链断裂,凋亡精子的比例显著上升[3]。苯并芘(BaP)皮下注射雄性小鼠时会促进小鼠精子、精母细胞凋亡,显著提高凋亡精子的百分比[45]。虽然目前细胞凋亡的分子机制尚未彻底明了,但已证实有多种基因参与该过程,如bcl-2基因家族、p53和Fas/FasL系统等。
2.2.1 通过干扰bcl-2诱导细胞凋亡
在细胞凋亡过程中,细胞色素C(Cyt C)和蛋白水解酶Caspase-3是重要的传递者和执行者,Cyt C携带着由线粒体发出的凋亡信号释放到胞浆中,募集此处未被激活Caspase,二者相互酶解活化,释放出有活性的Caspase,从而诱导细胞的凋亡[46-47]。bcl-2是B细胞淋巴瘤-2基因家族的成员之一,该基因编码的蛋白能够通过抑制线粒体中Cyt C的释放而阻断细胞凋亡的发生,但同属该基因家族的bax、bad和bak等基因所编码的蛋白可通过与Bcl-2形成异二聚体而抑制Bcl-2的功能,从而发挥着促凋亡作用[48-50]。而污染物可以通过改变Bcl-2、Bax和Bad的表达来促进细胞凋亡的发生。有研究表明,亚砷酸钠上调了雄鼠睾丸中促凋亡因子Bad的含量,下调了凋亡抑制因子Bcl-2的含量,增加了细胞质中Cyt C的水平,激活了Caspase-3的活性,最终导致生殖细胞凋亡率显著增加[15]。BPA通过诱导小鼠精原细胞Bax的高表达,下调Bcl-2的表达,使Bcl-2/Bax的比例失衡,显著提高Caspase-3的表达量,造成生精细胞凋亡比例增加,导致精子数量减少[51]。
2.2.2 激活p53途径介导细胞凋亡
抑癌基因p53也可诱发自发性、损伤性的生精细胞凋亡,它能够诱导线粒体结构损伤,促使线粒体释放凋亡诱导因子至胞质活化Caspase,从而执行细胞凋亡[52-53]。p53主要分布于雄性脊椎动物睾丸的初级精母细胞和间质细胞中[54],它在雄性生殖细胞的发育过程中,还具有监督作用,无论是外源性、内源性因素造成的精子发育异常,都会激活p53途径而促进异常细胞发生凋亡[55]。一些污染物可通过增强p53的表达而诱导细胞凋亡。如在金属锰染毒的大鼠的精原细胞和精母细胞中,p53蛋白的表达量显著升高,且这2种细胞的凋亡率也随之增加[52];壬基酚可通过上调大鼠睾丸中p53的mRNA及蛋白含量,显著上调caspase-1、-2、-9以及caspase-11的表达,导致小鼠精原细胞、初级精母细胞和精细胞凋亡率显著升高[53]。
2.2.3 激活Fas途径诱导细胞凋亡
细胞凋亡受体Fas主要位于细胞膜上,当其与特异性配体FasL结合后,会形成一个三聚体凋亡诱导复合物,该凋亡复合物中含有死亡结构域的Fas相关蛋白可以使Caspase活化,随后引起细胞凋亡[56]。雄性睾丸组织中的生精细胞、支持细胞和间质细胞均可表达Fas,该通路的激活是环境污染物诱导生殖细胞凋亡的主要方式之一。有研究表明,BPA可分别增加小鼠生精细胞和支持细胞中的Fas和FasL水平,活化生精细胞和间质细胞中的Caspase-3,从而促进了细胞凋亡[57]。而空气污染物PM2.5也可通过显著上调大鼠睾丸中Fas/FasL凋亡通路相关因子的表达,使Caspase-3/Caspase-8的表达显著增加,导致生殖细胞凋亡率升高,降低精子的数量[24]。
2.2.4 通过其他信号通路促进细胞凋亡
除上述经典的细胞凋亡信号通路外,核因子E2相关因子2(Nrf2)通路在调节细胞凋亡过程中也发挥着一定作用[58]。它可以与bcl-2启动子区的AREr3结合,来促进bcl-2的表达,Bcl-2增加后将抑制Bax、Cyt C释放和Caspase-3/7激活,从而抑制细胞凋亡[59]。若Nrf2通路受到干扰,细胞凋亡就会丧失一个有力的“阻碍者”,从而升高正常细胞的凋亡率。例如,雄性小鼠暴露于全氟烷基酸后,其睾丸中Nrf2的mRNA表达量及蛋白水平均显著下调,Caspase-3的含量显著升高,生殖细胞的凋亡数量显著增加[60]。另有研究表明,氟化钠也显著下调了大鼠睾丸中Nrf2的表达量,增加了Caspase-3的活性,提高了睾丸曲精小管中细胞的凋亡率[61]。
已有的研究报道,核因子kappa-B(NF-κB)通路对生精细胞的凋亡发挥着双向调控作用。一方面NF-κB能够激活凋亡抑制基因或蛋白(Bcl-xl、XIAP和Bcl-2)的表达而起到抗凋亡的作用[62-63];另一方面NF-κB能通过刺激Fas及Caspase的表达,促进细胞凋亡[64],NF-κB通路对细胞凋亡的总体调控效果取决于这2种作用之间的平衡。一些污染物可通过上调NF-κB促进细胞凋亡,如经林丹暴露后的大鼠,其睾丸生殖细胞中NF-κB的表达量明显增加,Fas的含量也相应上调,Caspase-3/9被活化,从而促进了生殖细胞凋亡[65]。此外,环境污染物还可以通过上调肿瘤坏死因子-α、白细胞介素激活NF-κB,来调节促凋亡因子的表达,从而促进细胞凋亡。如使用阿霉素注射雄性大鼠后,其睾丸中肿瘤坏死因子-α、白细胞介素-6的含量显著增加,NF-κB的含量相应升高,激活了Caspase-3,由此上调了睾丸中细胞的细胞凋亡率[66]。
3 改变雄激素水平(Change androgen levels)
精子发生和精子成熟是一个高度有序且受很多因素调控的过程。在雄性生物体内,下丘脑-垂体-性腺(HPG)轴是雄性类固醇激素合成的主要调控因素,而这些雄性类固醇激素又控制着精子的发生[67]。其中,雄激素睾酮(T)、二氢睾酮(DHT)和11-酮基睾酮(11-KT)在精子发生和精子成熟中具有关键作用[68-69]。T可以通过与支持细胞上的雄激素受体结合发挥作用,并在精子发生的特定时期刺激各种蛋白质和营养因子的合成[70],因此,T对减数分裂和随后的精细胞分化至关重要[71]。污染物会通过降低T的含量来影响精子的生成。研究发现,以60 μg·d-1剂量的BPA喂食小鼠,从交配前喂食至断奶,其雄性后代血中T的含量显著降低,并观察到生精小管以及精子数目减少[72]。
此外,DHT也可与雄激素受体结合而发挥相应的生理功能,研究发现,在人和大鼠中DHT可能与精子成熟有关,因为参与合成DHT关键酶5-α还原酶的活性变化与精子在附睾中成熟过程相一致[69];若抑制5-α还原酶的活性,则血清中DHT含量降低,精子数量显著减少[73]。而11-KT作为硬骨鱼类的主要雄激素,能够调节精原细胞的有丝分裂、精母细胞的形成和精子的生成[74]。11-KT可以诱导鳗鲡精原细胞进入减数分裂期,分化为可育的精子[75];其水平的降低与精子发生受阻和精子数量下降密切相关[76]。在环境污染物对雄性生物生殖影响的研究中发现,污染物可通过直接或间接2种方式扰乱雄激素的合成过程,如图2所示,从而影响生精过程。
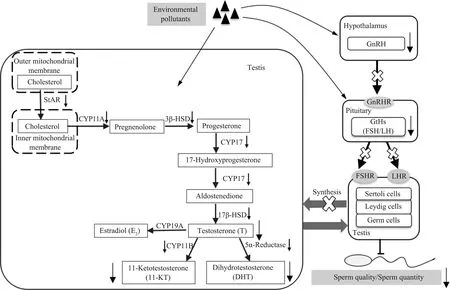
图2 环境污染物通过改变雄激素水平损伤精子的机制Fig. 2 The mechanisms of sperm damage induced by environmental pollutants via altering endogenous androgen levels
3.1 直接干扰雄激素合成过程
环境中的污染物能直接作用于雄性生殖器官,改变激素合成过程中的相关基因的表达水平,从而改变激素含量,最终影响精子的发生。例如,邻苯二甲酸二乙酯降低了雄性斑马鱼雄激素合成过程中相关基因cyp17a1、17βhsd的表达,从而使得血浆中T的含量明显下降,显著抑制了斑马鱼精子的形成[77]。经邻苯二甲酸二异丁酯暴露的小鼠附睾中参与雄激素合成的相关基因star、p450scc、cyp17a1、3βhsd和17βhsd的表达量显著下调,导致其血浆中T的含量显著下降,精子的活力及数量也随之降低[78]。使用除草剂利谷隆灌胃妊娠期大鼠后发现,子代雄性大鼠睾丸组织中参与T、DHT合成的star、p450scc、3βhsd和5α-R的表达量显著下调,且性成熟后子代雄鼠精液中的精子数量显著降低[79]。
有学者将成年雄性斑马鱼长期暴露于DES和氟他胺,发现合成11-KT的关键酶基因cyp11b2的表达水平显著降低,11-KT的含量也相应降低,进而阻断了精子发生[41];另一研究发现,除草剂阿特拉津能显著下调雄性斑马鱼精巢中cyp11b2的含量,使得11-KT水平随之降低,最终导致精巢中的精细胞密度显著下降[80]。可见,环境中的污染物可通过改变雄激素合成过程中相关基因及酶的表达水平,使得雄激素的水平发生异常变化,扰乱其所应发挥的生理作用,进而对生精过程产生不良影响。
3.2 通过调控HPG轴上游激素表达影响雄激素合成
下丘脑能合成并分泌促性腺激素释放激素(GnRH),GnRH会与位于垂体前叶促性腺激素细胞表面的GnRH受体结合,从而促进卵泡刺激素(FSH)和黄体生成素(LH)的产生[81]。而FSH和LH都是生理调控中非常重要的垂体激素,能以一种高度特异性的方式与其受体结合而发挥作用[82]。在脊椎动物中已经证实促性腺激素LH和FSH会参与调控精子的形成[83-84]。在精子发生过程中,LH会与间质细胞膜上的LH受体相作用,激活合成T过程中所涉及到的一系列CYP450酶和脱氢酶,刺激间质细胞从头合成T[85-86]。而FSH能与睾丸中支持细胞上的FSH受体结合,促进支持细胞合成并分泌雄激素结合蛋白,诱导精原细胞发生有丝分裂,促进精母细胞的发育,并与LH和雄激素协同作用促进精子发育成熟[87-88]。
污染物还会作用于中枢神经系统,通过改变GnRH、FSH和LH的水平来扰乱雄激素合成,进而影响生精过程。如全氟辛烷磺酸的暴露显著降低了雄鼠下丘脑中GnRH的含量,抑制LH的分泌,最终抑制T的合成,导致睾丸中的精子数量减少[89]。而汽油烟雾暴露后的小鼠,FSH和LH的含量被显著改变,血清中T的含量则显著降低,其精子数量和精子活力也表现出明显的下降[90]。因此,影响HPG轴上游激素的水平也会间接影响雄激素的合成,影响精子的发生。
4 其他可能的分子机制(Other possible molecular mechanisms)
精子发生是一个十分复杂的过程,环境污染物可能通过作用于其中任何一个靶点而对精子的数量和质量产生影响。在生精过程中,精原细胞是精子发生的基础,它们具有自我更新和分化成精子的能力[91]。精原细胞存在于干细胞生态位中,该生态位支持着精原干细胞最终分化为成熟的精子。该分化过程与α6-整联蛋白和β1-整联蛋白这2种细胞粘附分子的表达密切相关,并且这些整合蛋白在将精原细胞连接到它的生态位中起着至关重要的作用。研究发现,汽油尾气暴露会通过下调α6-整联蛋白和β1-整联蛋白导致雄性大鼠精原细胞受损,进而造成精原细胞、初级精母细胞、次级精母细胞和精细胞的总数显著减少,精子数量显著降低,精子畸形率明显增加[6]。
此外,在生物发育和分化的过程中,表观遗传学中对基因的活性和表达的调节是性腺正常发育和精子发生的基本机制之一[92]。异常的DNA甲基化模式可导致精子数量和质量的异常。而环境污染物会改变雄性生殖细胞的DNA甲基化水平,从而损伤精子。如镉可通过抑制DNA甲基转移酶的活性降低DNA甲基化水平,而基因组低甲基化会通过影响细胞的有丝分裂和减数分裂而干扰精子的正常形成,从而使成年雄性大鼠精子数下降[9,93]。并且这些表观遗传学的变化会遗传给子代,如有研究表明,啮齿动物在成年期间暴露于各种化学物质可以诱导其生殖细胞中DNA甲基化水平发生变化,导致遗传信息发生改变,这种改变会跨代传递并在后代中具有表型效应[94]。
近年来的研究表明,一些污染物如化学除藻剂新洁尔灭能够与鲱鱼精子的DNA以沟槽作用相结合[95];在吸烟男性的精子中发现BaP会与精子DNA形成BaP-二醇-环氧化物-DNA加合物[8],这种污染物直接与精子中生物大分子的结合,可能也是其损伤精子的原因之一。由此可见,环境污染物可以通过多种作用机制损伤精子数量及质量,进而对雄性生物的生殖及后代产生不利影响。
5 结语(Conclusions)
综上所述,环境中的污染物可以通过不同的作用方式损伤精子的形态结构、功能以及数目,导致精子质量和数量的下降。而精子质量受损、数量下降会造成生育力降低、繁殖障碍等多种生殖异常疾病,所以明确环境污染物损害精子的靶点和分子机制能够为预测污染物的生殖毒性效应提供依据,并为开发污染物对精子损伤的早期特异性分子标记物奠定理论基础。
此外,研究证实当亲本精子受损后会产生继代甚至跨世代影响。例如,父系精子基因组受到氧化性DNA损伤后,繁殖后的胚胎表现为DNA去甲基化损伤,发育停滞[96]。当使用氰戊菊酯灌胃成年雄性小鼠后,其精子畸形率显著增加,精子数量减少,生精细胞皆不同程度受损。同时,它与健康雌鼠繁殖后得到的子一代雄性小鼠在未受任何毒物暴露的情况下也出现了生精细胞结构异常,精子密度降低,精子畸形率升高,成熟精子染色质消失等现象,并且使用子一代雄鼠与健康雌鼠繁殖后得到的子二代小鼠的精子密度也显著降低[97],然而这种环境污染物造成的跨世代影响的分子机理和生理学机理尚不明确,可能成为今后环境污染物对精子损伤方面研究的一个新的方向。
猜你喜欢
右江医学(2021年9期)2021-10-22
昆明医科大学学报(2021年1期)2021-02-07
小学阅读指南·低年级版(2020年9期)2020-10-12
阅读与作文(小学低年级版)(2019年11期)2019-12-26
基础教育论坛·上旬(2019年7期)2019-09-10
中成药(2017年4期)2017-05-17
飞碟探索(2016年5期)2016-05-10
中国男科学杂志(2016年9期)2016-03-20
医学研究杂志(2015年12期)2015-06-10
中国当代医药(2015年1期)2015-03-01
