
泉州湾河口湿地互花米草种群的季节动态研究
2019-04-19 05:29高苏蕊李元跃陈融斌赵秋毅熊章静唐业勋黎中宝
集美大学学报(自然科学版) 2019年2期
高苏蕊,李元跃,陈融斌,潘 文,赵秋毅,熊章静,唐业勋,黎中宝
(1.集美大学水产学院,福建 厦门 361021;2.福建省海洋渔业资源与生态环境重点实验室,福建 厦门 361021;3.集美大学食品与生物工程学院,福建 厦门 361021)
0 引言
互花米草(Sporobolusalterniflorus(Loisel.) P.M.Peterson & Saarela)起源于美洲大西洋沿岸及墨西哥湾[1],由于人类有意或者无意的引入,现已在全球范围内成功存活、繁殖并迅速地传播蔓延[2],成为全球滨海湿地生态系统广泛分布的入侵植物之一[3]。由于互花米草的根系发达、繁殖力强,具有耐盐、耐潮汐淹没、对土壤适应性广等特点[4],被众多沿海国家所引种,用于保滩护堤、绿化海滩、改良土壤条件和改善海滩生态环境[5]。互花米草的生长速度远远超过人们的控制能力水平,因此在我国引种后呈现大规模爆发式增长,成为我国滨海湿地的外来入侵植物之一[6]。沿海滩涂湿地与河口湿地是单位面积上生态服务价值最高的两种生态系统类型,也是最容易被外来生物入侵的生态环境类型之一[7]。互花米草入侵导致滩涂泥沙沉降淤积,影响了滩涂养殖并阻塞航道[8],且互花米草的扩张挤压了其他潮间生物的生存空间[9],使湿地生物多样性发生了巨大改变,尤其是我国“海上森林”——红树林的生态系统结构和功能遭到了破坏[10],导致红树林严重的衰退和生物多样性的降低[11]。泉州湾河口湿地是中国亚热带河口滩涂湿地的典型代表,是重要的自然生态资源[12],2000年列入中国重要湿地名录,2003年建立泉州湾河口湿地省级自然保护区[13],它是生产力高、生态效应显著的区域,也是易受干扰、易发生变化的生境[14]。随着互花米草在该地大面积的入侵,泉州湾河口湿地面临的生态问题日趋加重。本研究以泉州湾河口湿地互花米草为研究对象,研究互花米草种群特征随季节和滩涂高程的动态变化,以期为泉州湾红树林湿地的保护和互花米草的治理提供参考。
1 材料与方法
1.1 试验地概况
本实验研究区域设在福建泉州湾洛阳江区红树林保护区的河口湿地(24°51′N~24°58′N、118°37′E~118°43′E)内。试验片区原是裸露滩涂,在互花米草成功入侵后成为互花米草单优草滩,其间仅发现少量人工种植的秋茄和桐花树等红树植物的存在。洛阳江是福建省泉州市的第二大河流,该地区为亚热带海洋性季风气候[14],年平均气温约20.2 ℃,极端最低气温和最高气温分别为0.1 ℃和38.7 ℃,全年降水时间主要集中在6~8月,是典型的河口湾和构造湾,水体主要受北支潮控制,属于规则半日潮,最大潮差为7 m[15]。
洛阳江区红树林保护区属于淤泥质砂质海岸、平原海岸、基岩海岸、局部红树林海岸的海岸类型,底质为粉砂质粘土至粗砂,有机物丰富,沉积层较厚。土壤类型为河口、滨海滩涂的淤泥和沙质淤泥[16]。该地区土壤理化性质为[17]:pH值为6.81~7.71(均值为7.12),盐度为0.92~1.76(均值为1.36),有机质含量为1.224%~2.041%(均值为1.538%),总氮含量为0.93~1.398 g/kg(均值为1.09 g/kg),全硫含量为0.031%~0.115%(均值为0.065%),碳氮比为6.880~9.447(均值为8.238)。
1.2 试验方案
分别于2017年6月(夏)、9月(秋)、12月(冬)和2018年3月(春)4次到泉州湾互花米草样地进行种群调查和采样。每次采样时,样地分高、中、低3个潮间带(垂直堤岸间大约间隔20 m)分别调查统计。每个潮间带断面随机选取3个1 m×1 m样方,统计样方内活的具有完整茎叶、株高高于地面10 cm以上的植株,记为1株。在这3个样方中选取6株植株带回实验室,洗净自然晾干测定株高、丛径、鲜重后送入烘箱105 ℃杀青1 h,再65 ℃烘至恒重后测定干重。
1.3 数据处理
数据统计分析软件为SPSS21.0,处理差异显著性分析采用S-N-K法检验比较(显著水平P=0.05),图表制作工具为Origin9.0。
2 结果
2.1 密度的季节动态变化
经样地现场调查,泉州湾互花米草各个生长阶段对应的时间为:3月份进入小苗的生长时期;6月份互花米草进入营养生长期;7—8月份互花米草已经开始进入生殖生长的扬花期;9月份以后互花米草生殖生长基本完成并开始逐渐大面积变黄枯萎,除极小面积倒伏外基本上仍竖立不倒,直到第二年新苗破土之前,枯死的互花米草茎杆开始大面积倒伏,为新苗腾出空间和阳光等养分。泉州湾互花米草植株密度随季节变化的结果见表1。

表1 不同潮位互花米草种群密度的季节变化
说明:表内数据为平均值±标准差 ;同列上标小写字母不同表示存在显著差异(P<0.05)
Notes:Data in the table is mean±SE,different lowercase letter in the same column indicate statistically significant differences(P<0.05)
从表2可以看出,随着季节的变换,互花米草从生长旺盛期到新旧交替期,三个潮区互花米草种群密度都是呈现夏季>秋季>春季>冬季。秋季和春季各个潮区互花米草的种群密度基本上一致,夏季和冬季各个潮区互花米草的种群密度则有较大差别,且冬季的种群密度随高程的上升而呈上升趋势。
经双因素方差分析得,季节变化和潮位均对互花米草的种群密度变化具有显著影响(P<0.01),且不同潮区四个季节之间互花米草植株密度变化差异极显著(P<0.001)。互花米草种群密度夏季显著高于秋季,秋季显著高于春季,且春季显著高于冬季 (P<0.01)。潮位低部互花米草的种群密度与潮位中部、高部之间差异极显著(P<0.001),潮位高部和中部互花米草的种群密度差异显著(P=0.037<0.05)。夏季潮位中部互花米草密度显著高于潮位高部和低部,冬季潮位高部互花米草密度显著高于潮位中部和低部。
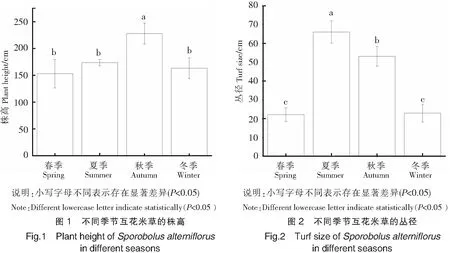
2.2 株高的季节动态变化
株高反映了植物地面部分垂直方向生长的情况。本研究中互花米草全年株高153.00~228.00 cm。由图1可知,互花米草的株高在秋季显著高于其他季节,其他季节株高差异不显著。在四个季节中互花米草的株高呈现秋季>夏季>冬季>春季。其中,夏季到秋季显著增长54.35 cm(增长率为31.30%),秋季到冬季株高显著降低64.85 cm,在秋季株高达到最大值(228.0019.49)cm。
2.3 丛径的季节动态变化
丛径反映了植物地面部分水平方向生长的情况。本研究中互花米草全年丛径为22.09~66.04 cm。由图2可知,从春季到冬季丛径的变化呈先上升后下降的趋势,且随季节变化差异均显著(P<0.05)。其中,春季到夏季增长43.95 cm,夏季到秋季与秋季到冬季分别显著减少12.90 cm和30.24 cm,在夏季达到最大值(66.04±5.91)cm。
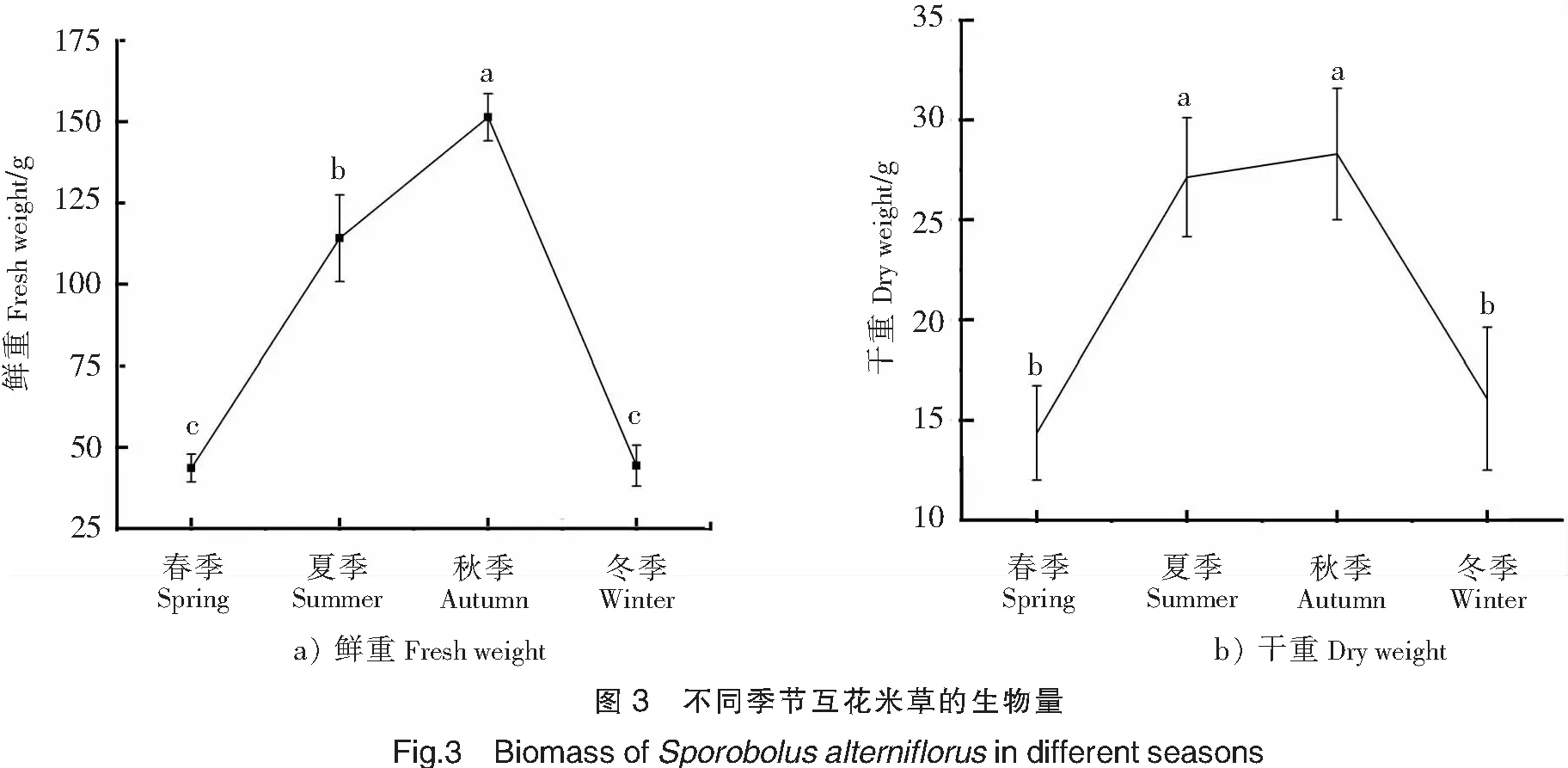
2.4 鲜重与干重的季节动态变化
鲜重与干重反映了种群的生物量。本研究中互花米草全年干重14.37~28.30 g。由图3可知,互花米草的鲜重与干重均在秋季较高,春季和冬季较低。在四个季节中互花米草的鲜重和干重均呈现秋季>夏季>冬季>春季。互花米草的鲜重和干重均在秋季达到最大值,分别为(151.417.24)g和(28.303.29)g。鲜重随季节变化差异均显著(P<0.05);干重在春季到夏季增加显著(P<0.05),增加了12.77 g,在秋季到冬季显著减少了12.23 g。
3 讨论
在海岸盐沼生境中,高程及其决定的水位是影响植物在滩涂上分布的重要生态因子,水位条件的变化可以改变植物在种间竞争中的相对表现,进而影响其空间分布[18]。黄冠闽[19]研究表明,盐度对互花米草的生长有显著的抑制作用,随着盐度的增加,互花米草的株高、基径、叶面积都显著降低。苑泽宁[20]研究发现,互花米草对盐度有较强的适应性,盐沼生境对植物生长的影响主要表现在高盐浓度介质造成的高渗透压胁迫,互花米草的结构与生理生化特征使其对高盐浓度具有一定的耐受能力。张亦默等[18]研究发现,互花米草对潮汐带来的淹水具有良好的适应能力,由于其具有发达的通气组织,可以提高其根周围土壤的溶氧度,因而其根系可以进行正常呼吸并吸收养分,能适应低潮区生存。季节引起的温度、光照、湿度等因素的变化,也直接对互花米草的种群特征产生直接的影响。
在不同地域互花米草的对比研究中,段博文[21]测定了湄州湾互花米草全年密度为57~107株/m2,株高为76.08~179.98 cm,丛径范围为13.63~53.07 cm,干重为8.19~27.29 g。与之相比,泉州湾河口湿地互花米草在各潮位的密度、株高、丛径、鲜重和干重等生物量均高于湄州湾的互花米草。经分析判断,互花米草生长特征随纬度变化会呈现明显二次曲线趋势[3]。泉州湾河口湿地位于湄洲湾的南部,所处纬度位置更适合互花米草生长;并且泉州湾滨海湿地为淤泥质砂质海岸且底质为粉砂质粘土至粗砂,有机物丰富,沉积层较厚[6],与湄洲湾沙质的底质相比更适于互花米草的营养繁殖;再者,有研究结果表明互花米草的最适生长盐度为10~20[22],超过该范围时,互花米草的生长会受到抑制,而湄洲湾的土壤盐度为3.5~28.9,相比于泉州湾盐度可能会抑制当地互花米草的生长。
在比较各季节不同潮区互花米草密度时发现,泉州湾不同潮区互花米草的密度均是夏季最高,冬季最低,且夏季到秋季密度变化幅度最大。在夏季中不同潮区密度差异显著,且表现为中潮区密度最高,可能与其盐度与水淹时间更适于互花米草的生长与扩散有关。冬季高潮区互花米草密度高,主要原因是低潮区和中潮区新苗出现时间比高潮区早,枯杆开始倒伏,腾出了新的空间,而高潮区受潮水淹没时间短且还未出现新苗,发达的根系也改善了土壤内部的环境,汲取土壤养分能力高于中低潮区,所以高潮区互花米草密度要大于低中潮区。
经数据分析:泉州湾互花米草性成熟约在6—8月。在春季进入返青期,对水分和营养成分的吸收开始恢复;夏季是互花米草快速生长时期,它能充分地利用阳光、土壤养分、生长空间,因为互花米草种群生长密集,如不能充分利用有限空间,则互花米草繁殖体就会变少,扩散能力大大减弱;在秋季互花米草生殖生长基本完成,由于气候及自身代谢等原因互花米草繁殖后开始进入黄枯期,这个时期持续时间较长,在这个时期里,枯萎的互花米草依然竖立,其一个原因可能是因为互花米草具有粗壮的茎杆,能强有力地支撑整个植株,另外一个原因可能是互花米草的根系发达,其庞大的根系能牢牢地固定地上部分;冬季由于气温和光照等因素的不利影响,互花米草枯杆逐渐开始倒伏,株高、丛径和鲜重等生物量均处于一年中较低。从上面分析可以判断,互花米草分蘖和生长时期在春季与夏季之交,生物量增速最快,是扩散能力最强的季节,而到了冬季,互花米草开始枯萎和倒伏,生物量相对较小,其扩散能力相对较低。从防控互花米草的角度看,在春季进行人工防控互花米草,可以大量减少互花米草的扩散,除此之外,由于春季的互花米草刚开始分蘖生长,其茎秆比较稚嫩,人工刈割比较容易,可以大量减少人力,节约人工成本。因此,从防控效果和成本的角度出发,在春季进行人工防控互花米草是最理想的。
猜你喜欢
农业灾害研究(2022年7期)2022-09-08
武汉理工大学学报(交通科学与工程版)(2022年4期)2022-09-07
安徽农业科学(2022年14期)2022-08-04
港工技术(2022年2期)2022-05-12
海洋通报(2021年5期)2021-12-21
河北地质(2021年1期)2021-07-21
能源与环境(2020年4期)2020-09-02
农村百事通(2019年17期)2019-10-08
水利水电快报(2019年2期)2019-03-08
现代农业科技(2017年1期)2017-03-06
