
遮荫对刈割互花米草生物量及渗透调节物质的影响
2019-07-03 09:17张丽平周亚圣刘君成尤丽萍韧蔡金标游巍斌何东进
西南林业大学学报 2019年4期
张丽平 周亚圣 刘君成 尤丽萍 王 韧蔡金标 游巍斌 何东进,5
( 1. 福建农林大学林学院,福建 福州 350002;2. 福建农林大学海洋研究中心,福建 福州 350002;3. 福鼎市林业局,福建 宁德 355200;4. 霞浦县林业局,福建 宁德 355100;5. 福建农林大学金山学院,福建 福州 350002)
互花米草(Spartina alterniflora)原分布于北美洲中纬度大西洋海岸潮间带滩涂,属于生命力极强的多年生耐盐耐淹米草属禾本科植物, 具有强大的克隆整合与环境饰变能力[1-2],能在沿海地区的热带、亚热带以及温带分布,通常在沿海滩涂的潮间带形成单优势群落[3]。我国闽东滨海地区出于保滩护堤的目的,于20世纪80年代初开始引入互花米草,如今已有近40年的历史,由于其繁殖扩散能力极强,引种成功后严重危害当地的生态环境[4-5]。互花米草作为一种喜光的C4植物,采取遮荫胁迫的手段可以极大程度上遏制其地上及地下生物量的生长。据苗圃模拟试验结果发现,在3层遮荫网遮荫处理下,互花米草植株1个月内即可全部死亡[6]。赵相建等[7]研究发现通过刈割加不同遮荫率的集成方法能够极大程度上抑制该植物地上生物量的生长。物理法中的刈割手段一方面可以抑制互花米草生长速度并降低其地上生物量,从而极大降低了光合产物向地下输送,影响根系生长和地下部分幼芽的再次萌发;另一方面通过阻断大气中的氧气通过茎器官等往地下传递,从而削弱其根部的有氧呼吸[8]。可见,植物对刈割和遮荫抗性并不是单一的表现,在逆境胁迫下,植被的种种形态数量特征的变化都可以在生理生化过程的改变中得以体现。
近年来,国内外学者对入侵种互花米草已展开了防控治理、生长生理机制响应、与红树林的竞争效应[9]、生态风险评价与预警[10]、土壤生物群落特征[11]等方面的研究,尤其在防控治理方面研究颇多,如通过物理、化学、生物控制和综合防治法等手段来遏制互花米草的生长,从而将其彻底根除[12]。以上关于互花米草的各领域研究颇多,但目前关于在霞浦闽东滨海湿地通过研究刈割加不同水平遮荫胁迫对互花米草地上生物量及渗透调节物质的影响未见研究。本研究以外来入侵种互花米草幼苗为试验材料,在闽东滨海湿地开展了刈割加不同遮荫处理试验,以单纯刈割(无遮荫)为CK,分析在不同遮荫率互花米草生物量以及渗透调节物质的变化规律,为揭示或找出该入侵种治理的最佳遮荫环境提供数据支撑。
1 材料和方法
1.1 试验区概况
本研究区位于霞浦县牙城镇杨家溪村,地处东经 119°46′~120°26′,北纬 26°25′~27°9′。霞浦县为福建省宁德市下辖的1个县,年平均气温为16~19 ℃,春季雨水丰沛,夏季台风多发,冬季温暖,霜雪天气少见。受海洋气候影响大,季风性气候特点十分明显,灾害性天气频繁,常以台风、暴雨为主,影响较大的台风年平均出现3次,属典型的中亚热带海洋性季风气候[13]。年平均降水量1 100~1 800 mm,年平均日照时数达1 637.7 h,境内平均海拔总体低于1 000 m,其中以目海峰海拔最高,约1 192.4 m,历史上曾先后2次出现极端气温,分别是1989年的极端最高气温39.1 ℃和1962年的极端最低气温-3.4 ℃。陆地和海域面积分别为1 489.6 km2、29 592.6 km2,此外,闽东滨海地区霞浦县处于中国南北海岸线的中心位置,南接珠江三角洲,北连长江三角洲,东与台湾岛隔海相望,海岸线约480 km,浅海滩涂面积约693.34 km2,大小岛屿共计196个,均居福建省各沿海县(市)的首位[14]。
1.2 试验方法
1.2.1 样地设置与采样
分别于2018年7月18日、8月20日、9月21日、10月22日在霞浦县牙城镇杨家溪村互花米草分布密集区先后进行4次采样,并选取15块4 m×5 m的样地,共计300 m2。据调查该时期(7月18日)植株地上部分平均高度为168.32~175.19 cm,平均密度112株/m2,然后将15块样地分布的互花米草全部人工刈割,整个试验期间每块样地刈割1次,刈割高度距离地面5~10 cm,并在此基础上分别用30%、50%、70%和90%遮荫网对其中12块样地进行覆盖处理,每处理3次重复,剩余3块样地不进行遮荫,作为CK。最后,每块试验地选取3块1 m×1 m的小样方,其间距确保大于1 m,每块小样方内随机采集5株高度、长势基本一致的互花米草植株装入自封袋,带回实验室进行生物量、生理指标(摘取靠近顶端的2~3片新鲜叶子)的测定。
1.2.2 指标测定
先将野外采集的互花米草进行叶与茎的分离,洗净分装后放至鼓风干燥箱(温度设为85 ℃)烘干至恒质量,利用高精度电子天平称重仪(精确到0.000 1 g)对其准确称量,记录干质量数据并计算生物量。参照参考文献[15] 测定互花米草渗透调节物质参数,每个指标重复测定3次。
1.3 数据分析
运用Microsoft Excel 2007软件进行数据处理与制图,采用SPSS 21.0软件进行数据统计分析与检验,并利用最小显著差异法(LSD)检验不同处理组间样本的显著性。
2 结果与分析
2.1 遮荫对刈割互花米草生物量的影响
由表1可知,遮荫前(7月份)互花米草叶生物量、茎生物量、样方内生物量在不同遮荫胁迫组间并没有表现出显著差异。但之后的3个月随着时间的延长和遮荫强度的加大,各遮荫处理互花米草叶、茎、样方内生物量总体上表现出下降趋势,且均低于同时期的CK,各遮荫处理除了30%遮荫处理外,剩余组均通过了差异性检验,与CK存在显著差异(P<0.05)。其中,尤以90%遮荫下的互花米草生物量与CK差异最大,与遮荫前相比,在遮荫期满时(3个月后),90%遮荫下的互花米草叶、茎以及样方内生物量分别为 2.12、3.64、474.83 g,分别只占同期 CK的43.35%、39.95%和45.16%。此外,从对互花米草各部分生物量的抑制效果分析,不同遮荫处理可排序为90%遮荫>70%遮荫>50%遮荫>30%遮荫>CK。可见,高强度的遮荫处理环境可以极大程度限制互花米草地上生物量的增长。尽管高强度遮荫可以极大降低互花米草的生物量,但是在野外发现不同遮荫处理下互花米草新生苗也在不断萌发,各遮荫处理并没有彻底清除或消灭该入侵种地上部分的植株,特别是低强度遮荫环境下互花米草新生植株萌发率高。因此,可以推断高强度的遮荫加上更长的治理时间在根除互花米草复萌的效果上更加明显。

表 1 不同遮荫处理对互花米草生物量的影响Table 1 Effects of different shading treatments on biomass of S. alterniflora
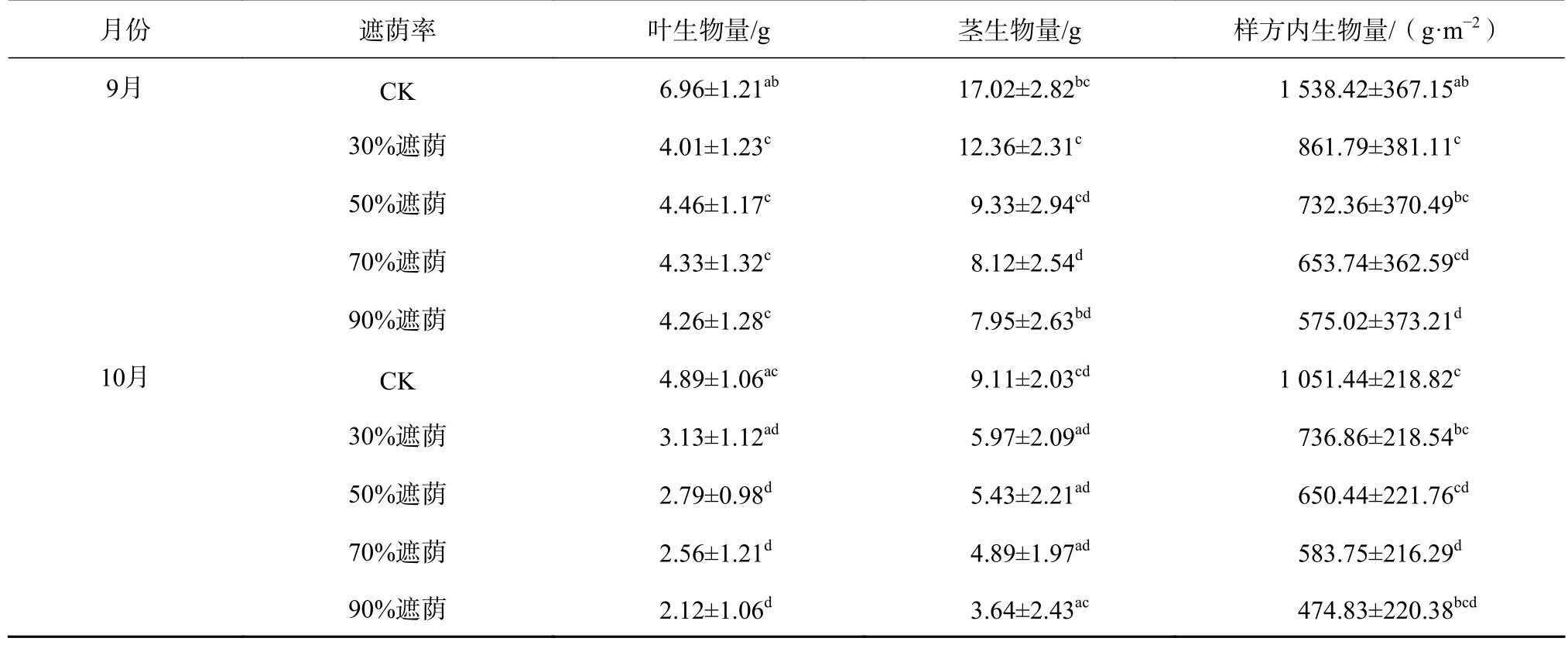
续表 1
2.2 遮荫对刈割互花米草游离脯氨酸含量的影响
不同遮荫处理下互花米草叶片游离脯氨酸含量的变化见图1。在整个遮荫期间,随着遮荫强度的加大,互花米草叶片游离脯氨酸含量在不断增加,其中CK中的游离脯氨酸在遮荫处理3个月后达到峰值,由起初的 60.55 μg/g上升到101.61 μg/g,其他处理遮荫中游离脯氨酸含量波动最大的是90%遮荫处理,波动为61.37~241.59 μg/g,波动最小的是30%遮荫处理,波动为58.42~119.49 μg/g。同一遮荫下随着时间的延长,互花米草游离脯氨酸含量除CK不断上升外,其余各组均表现出先升后降的单峰变化趋势,互花米草叶片游离脯氨酸含量在不同遮荫处理下出现这种变化趋势,这可能与其适应性调节机制有关。除30%遮荫处理外,其他遮荫处理互花米草叶片游离脯氨酸含量与同期CK相比均存在显著差异(P<0.05)。
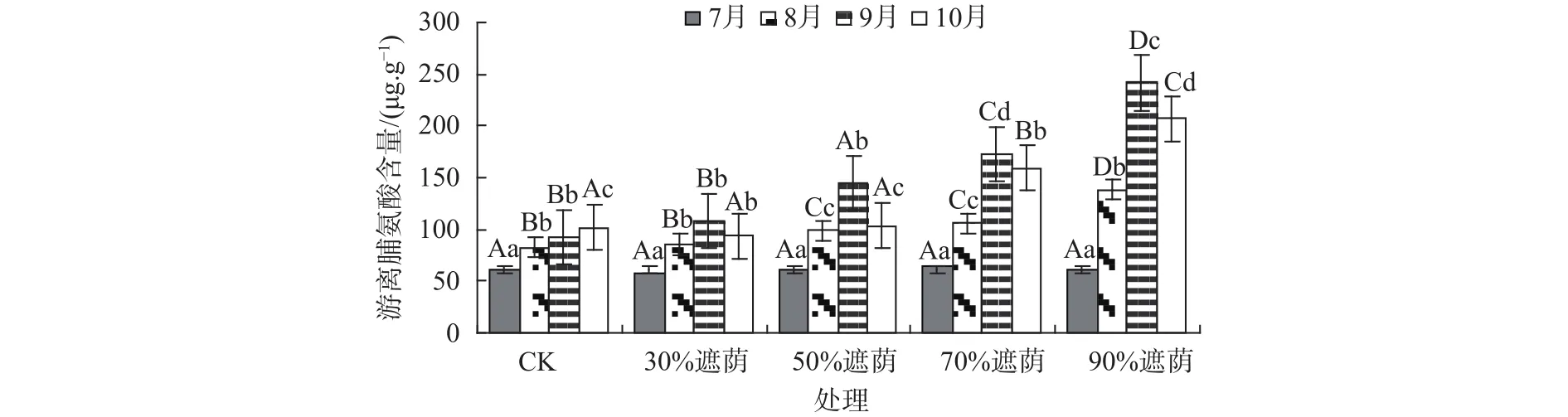
图 1 不同遮荫处理下互花米草叶片游离脯氨酸含量Fig. 1 Content of free proline in leaves of S. alterniflora under different shading treatment
2.3 遮荫对刈割互花米草可溶性糖含量的影响
由图2可知,在不同处理下,随着遮荫强度的变大,各遮荫处理互花米草叶片可溶性糖总体上均呈下降的趋势,但在遮荫末月其含量呈现增高的趋势,遮荫强度越大,互花米草可溶性糖含量增幅也越大,这说明在高强度遮荫环境胁迫下互花米草可溶性糖含量响应要比低强度遮荫环境胁迫积极。在遮荫期间,CK的该米草叶片可溶性糖由起初的45.66 mg/g降到22.84 mg/g,其他处理中可溶性糖含量波动最大的是90%遮荫处理,波动为9.89~46.03 mg/g,波动最小的是30%遮荫处理,其可溶性糖含量为24.69~44.87 mg/g。而在同一处理下互花米草叶片可溶性糖含量,除CK不断下降外,其余各组均随胁迫时间的增加表现出先降后升的趋势,这种变化趋势性可能与其适应性调节机制有关。除30%遮荫处理外,其他遮荫处理的互花米草叶片可溶性糖含量与同期CK相比均存在显著差异(P<0.05)。
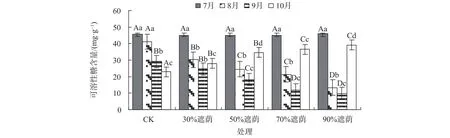
图 2 不同遮荫处理下互花米草叶片可溶性糖含量Fig. 2 Content of soluble sugar in leaves of S. alterniflora under different shading treatment
2.4 遮荫对刈割互花米草可溶性蛋白含量的影响
由图3可知,在各遮荫环境下,随着遮荫强度的逐渐增强,互花米草可溶性蛋白含量在不断下降,且遮光率的强度越大,可溶性蛋白含量的变幅也越大,可见,在高强度遮荫下互花米草叶片可溶性蛋白响应要比低强度遮荫胁迫下积极。在遮荫期间,CK的互花米草可溶性蛋白含量由起初的82.39 mg/g降到46.81 mg/g,其他处理中可溶性蛋白含量波动最大的仍是90%遮荫处理,为15.48~81.14 mg/g,波动最小的是30%遮荫处理,为37.46~83.25 mg/g。同一遮荫环境下互花米草可溶性蛋白除CK先略微升后降外,其余各组均表现出波动下降的变化趋势,互花米草叶片可溶性蛋白含量在不同遮荫处理下出现这种变化规律,这可能与其适应性反馈效应调节机制有关。除30%遮荫处理外,其他遮荫处理的互花米草叶片可溶性蛋白含量与同期CK相比也均显著差异(P<0.05)。
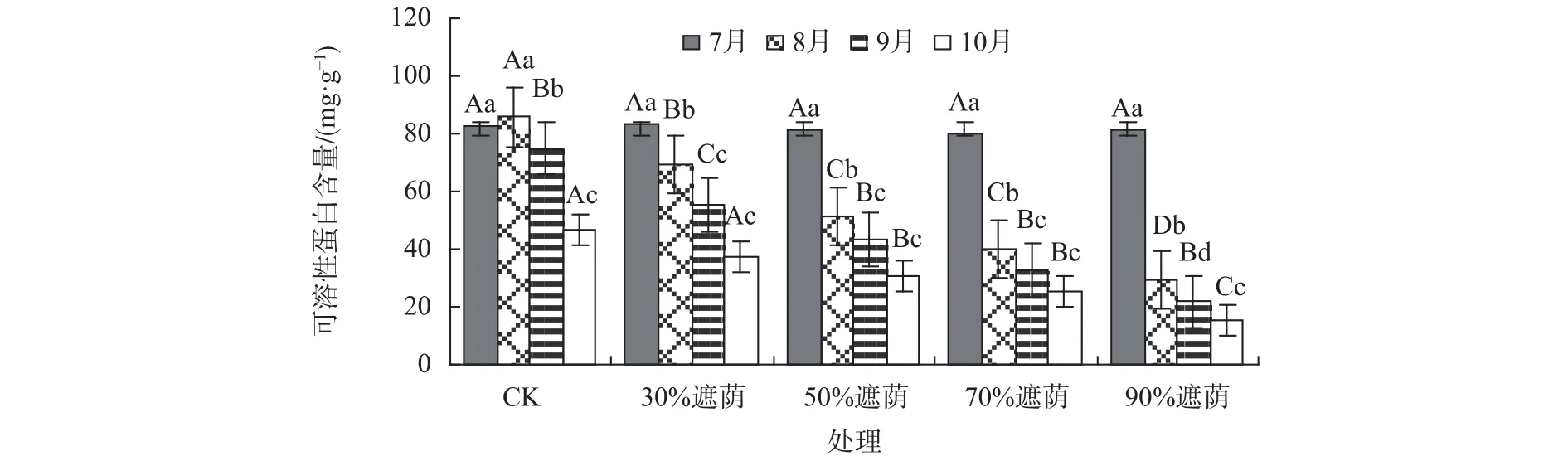
图 3 不同遮荫处理下互花米草叶片可溶性蛋白含量Fig. 3 Content of soluble protein in leaves of S. alterniflora f under different shading treatment
3 结论与讨论
外界环境因子(比如干旱、光照、水分、气温、稀土元素等)胁迫会引起植物生长及生理特性的一系列变化,是限制植物生长的重要因素[16-17],在暗环境(低透光率)下植物生长会受到抑制,光合效率明显下降,生物量、可溶性糖和可溶性蛋白含量急剧减少,最后导致植物叶片减少、枯黄、凋落甚至死亡[18]。本研究于2018年7月份在霞浦县杨家溪村开展刈割加不同水平遮荫试验,在刈割加不同程度遮荫试验的随后3个月互花米草叶生物量、茎生物量以及样方内生物量均呈现大幅下降趋势,很大程度上抑制了互花米草的生长,这与赵相建等[7]、张炯森[19]的研究结果一致,但在互花米草生物量清除力度上与他们的研究结果存在差异。本研究所设置的各遮荫处理并没有彻底清除互花米草地上部分的植株,究其原因很可能在遮荫时间和遮荫方式上与两者有区别,本试验遮荫胁迫时间仅3个月,并且遮荫网与地面存在1.4 m左右的高度,这就使得互花米草生存空间较大,再加上胁迫时间较短,从而无法彻底清除互花米草地上部分,抑制效果不佳,而赵相建等[7]、张炯森[19]均用贴地覆盖的方式对互花米草进行治理,这极大缩小了互花米草的生存空间,外加遮荫时间较长,因此在防控、清除该米草效果上更加明显。尽管三者的研究在互花米草防控效果上存在差异,但都说明了高强度遮荫环境对互花米草的抑制效果更显著,可以极大降低其地上部分生物量。
此外,本研究结果显示,不同遮荫处理对互花米草的渗透调节物质有显著的影响,随着遮荫强度的加大,互花米草可溶性蛋白和可溶性糖在不断减少,一直处于不断消耗状态,可溶性糖、可溶性蛋白含量不断降低的原因可能是在低透光率环境下互花米草光合作用受到限制,在这种环境下为了获取生存所需要的能量,从而消耗了植物体内大量碳水化合物以及可溶性糖,使得植物体糖分剧烈消耗,出现“糖饥饿”现象[20],植物可通过渗透调节物质脯氨酸来调节自身细胞渗透势,在保护蛋白分子与相关的酶活性方面作用突出,其含量的增高可能是自身抵抗外界渗透胁迫的一种抗逆境机理[15]。随着遮光程度的加大,游离脯氨酸不断积累增加,该变化规律可被解释为互花米草对不同遮荫环境下的一种抗逆机制做出的响应。此外,不同透光度环境下对植物的生长和生理影响也存在差异性,在胁迫期间,各遮荫处理除了30%遮荫处理外,其他遮荫处理互花米草生物量、渗透调节物质与同期CK相比均通过了显著检验,存在显著差异,尤以90%遮荫处理差别最大,这与王凯[21]的研究结论吻合,说明高强度遮荫对互花米草生物量和渗透调节物质影响更为显著,可以极大抑制该入侵种的快速生长。
综上所述,虽然单纯刈割可以快速显著的降低互花米草株高,从而减少其地上部分总生物量[22],对其生长和生理有一定程度的抑制效果,但随着时间延长,单纯刈割治理成效不佳,互花米草很快又恢复到治理前的水平,说明单一刈割只能在短时间内效果显著[23]。而本研究发现刈割加高强度遮荫环境(90%遮荫率)对互花米草生物量和渗透调节物质影响相比其他处理组更为显著,可以极大程度上抑制互花米草的生长,显著减少其地上部分的生物量,不失为一种可取的防控手段,值得推广。
猜你喜欢
中老年保健(2022年3期)2022-11-21
农业灾害研究(2022年7期)2022-09-08
安徽农业科学(2022年14期)2022-08-04
海洋通报(2021年5期)2021-12-21
今日农业(2020年19期)2020-12-14
能源与环境(2020年4期)2020-09-02
棉花学报(2020年3期)2020-08-08
科学与财富(2019年15期)2019-10-21
中国科技信息(2015年2期)2015-11-16
植物营养与肥料学报(2014年1期)2014-03-11
