
福建省永安市丝栗栲次生林林分结构特征研究
2019-08-20 14:58罗洁明张巧巧常赜陈昌雄
江苏农业科学 2019年8期
关键词:空间分布
罗洁明 张巧巧 常赜 陈昌雄


摘要:在丝栗栲次生林的阳坡和阴坡各设置1块50 m×50 m的典型样地,样地中丝栗栲种群占绝对优势,丝栗栲幼苗幼树占林分更新总株数的32.29%。阳坡丝栗栲种群密度和幼苗幼树(DBH<7.5 cm)密度均小于阴坡,但中树(7.5 cm≤DBH<22.5 cm)和大树(DBH≥22.5 cm)密度大于阴坡。2个坡向均表现为幼苗幼树多,中树大树少,属于增长型种群,幼苗贮备量大。2个坡向幼苗幼树密度在地径和树高等级上差异不明显,随地径和树高的增大整体呈递减趋势。地径分布呈倒“J”形曲线,树高分布呈明显的非对称性山状曲线,接近幼苗幼树平均高的株数较多,幼苗幼树地径和树高结构良好,供给稳定。
关键词:丝栗栲;种群结构;空间分布;福建省永安市
中图分类号:S718.5 文献标志码: A
文章编号:1002-1302(2019)08-0162-03
丝栗栲是福建常绿阔叶林生态系统的重要组成部分,经济、社会及生态效益显著,科学研究价值较高。但现存的丝栗栲林多为次生林[1],天然更新速度减慢,分布面积也在不断减少,使得其较强的培肥土壤、涵养水源能力无法充分发挥。福建省永安市的丝栗栲林分多为低产次生林,深入探索丝栗栲次生林中不同坡向絲栗栲种群及更新幼树幼苗在林分结构上的特有规律,对于揭示种群更新具有重要意义,可以促进该地区丝栗栲次生林经营管理与可持续利用,维持区域内的生态平衡。
1研究地概况[LL]
福建省永安市(116°56′~117°47′E,25°33′~26°12′N)位于福建省中部偏西,总面积2 942 km2,其中林业用地25.5万hm2。该市属典型的亚热带湿润季风气候,年均气温14.3~19.2 ℃,年降水量1 490~2 060 mm[1]。土壤以红壤和黄壤为主[2],植被属于中亚热带照叶林植被带,植被类型多样[3]。
2研究材料与方法
2.1样地设置
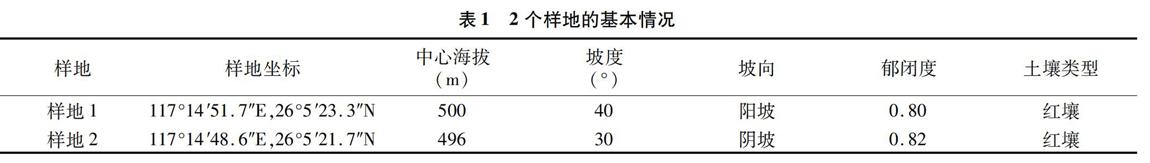
在丝栗栲次生林的阳坡和阴坡各设置1块50 m×50 m的典型样地,并进行每木检尺。样地基本概况如表1所示。
2.2林分结构分析方法
植物个体的径级和高度能够有效反映种群现状和发展趋势[4]。野外调查时,测定每个种群个体的年龄较为困难,种群龄级和径级对同一环境的反映有相对一致性[5],所以在前人研究[6-7]的基础上,可以利用立木级代替年龄来研究种群的年龄结构[8]。
根据树木的胸径(diameter at breast height,DBH)大小,将丝栗栲活立木划分为4个发育阶段。Ⅰ级:DBH<2.5 cm为幼苗;Ⅱ级:2.5 cm≤DBH<7.5 cm为幼树;Ⅲ级:7.5 cm≤DBH<22.5 cm为中树;Ⅳ级:DBH≥22.5 cm为大树[9-10]。
根据地径(diameter at ground height,DGH)大小,对丝栗栲种群天然更新幼苗幼树的地径进行以10 mm为标准的等级划分。一级(Ⅰ):0 mm 根据树高(height,H)的大小,对幼苗幼树进行高度级划分。一级(Ⅰ):H≤0.33 m;二级(Ⅱ):0.33 m 2.3偏度、峰度和变动系数 采用偏度、峰度和变动系数3个指标[11-12]对丝栗栲种群直径分布特征进行描述。 偏度:Cs=n(n-1)(n-2)∑ni=1xi-xs3;(1) 峰值:K=[JB({]n(n-1)(n-2)(n-3)∑ni=1xi-x[TX-]s4[JB)}]=3(n-1)2(n-2)(n-3);(2) 变动系数:C=sx。(3) 式中:Cs是衡量非对称分布偏斜方向与偏斜程度的指标,Cs>0表示左偏(正偏),均值在峰值左边;Cs<0表示右偏,均值在峰值右边;Cs=0表示对称分布;Cs绝对值越大,则表明偏斜程度越大。K是衡量曲线分布尖峭或平坦程度的指标,K>0表示尖峭,K<0表示平坦,K=0为正态分布;K值愈大,表明曲线愈尖峭,反之愈平坦。C反映分布范围的大小,其值愈大,表明分布离散程度愈大。xi为第i株的直径,x为平均直径,n为总株数,s为标准差。 3结果与分析 3.1丝栗栲种群结构特征 根据调查结果统计丝栗栲种群不同坡向两个样地内幼苗、幼树、中树、大树4个立木级的株数分布情况(表2)。从表2可见,在两个样地内Ⅰ级立木最多,依次减少,Ⅳ级立木最少。其中,相对于样地2,样地1中Ⅰ级立木占的比例有所下降(约10%),Ⅱ级立木的比例相差不大,而Ⅲ级立木的比例有所提高(约7%)。样地2幼苗和幼树的累计比大于样地1,中树和大树的累计比小于样地1。 直径分布特征值见表3。2块样地的种群林木直径标准差在7 cm以上,表明2块样地的直径变动较大,样地1的标准差大于样地2,说明样地1的直径变动范围更大,但2个样地的变动系数(C)相差不大,说明2个样地的直径变动程度相近。从偏度系数(Cs)可以看出,2块样地的Cs值均大于0,表明2块样地的林木直径分布曲线均为左偏曲线,而且样地2的左偏斜程度大于样地1,说明样地2的直径分布更偏向于中小径木。从峰度系数(K)来看,2块样地均为负数,表明直径分布曲线比较平坦,直径分布的离散程度较大,这可能与种群密度、平均直径以及生境条件综合作用有关。在整个丝栗栲次生林林分中,丝栗栲小径阶株数占种群的比重最大,株数随径级的增大而递减,达到一定径阶后,递减速度变小最终达到稳定状态。以上分析表明丝栗栲种群直径分布为近似倒“J”形。 3.2丝栗栲幼苗幼树的地径和树高结构 由图1可知,丝栗栲种群更新苗在样地1和样地2表现不同,在样地1中Ⅱ级幼苗数量最多,Ⅰ级次之,Ⅶ、Ⅷ级最少,径级结构分布曲线呈单峰左偏状;在样地2中Ⅰ级最多,Ⅱ级次之,Ⅷ级最少,径级结构分布曲线基本呈倒“J”形。样地2的Ⅰ~Ⅱ级更新苗数量(1 136株/hm2)比样地1(660株/hm2)多72.12%。丝栗栲幼苗具有耐阴性,阴坡较弱光环境更适合丝栗栲幼苗生长。另外,样地1中Ⅴ和Ⅶ级幼树分别为样地2的57.14%、41.67%,而Ⅳ级(132株/hm2)却明显高于样地2。综合来看,次生林中丝栗栲更新幼苗幼树株数随地径级的增加呈降低趋势,分布曲线呈倒“J”形,幼苗幼树结构为增长型。 丝栗栲更新幼树幼苗的高度结构分布见图2。样地1中,更新苗在Ⅰ~Ⅵ高度级的株数分布差异不明显,占总株数的91.76%。样地2中,Ⅱ级株数最多(464株/hm2),其次是Ⅰ级(292株/hm2),在Ⅳ级(248株/hm2)和Ⅶ级(144株/hm2)出现2个次高峰。从图2可以看出,样地1的更新高度结构曲线呈明显的左偏山状分布,而样地2呈近似双峰型分布。整体来看, 丝栗栲幼树幼苗在次生林中呈明显的非对称性山状曲线,接近幼苗幼树平均高的株数较多,Ⅱ~Ⅳ级占总株数的53.04%,Ⅶ、Ⅷ级株数较少。 3.3丝栗栲更新幼苗幼树地径和树高的空间分布 为清楚、明了地看出样地内丝栗栲更新苗木的空间分布情况以及地径和高度结构的空间分布情况,分别绘制了更新幼苗幼树的地径和树高空间分布图(图3和图4)。 样地1中幼苗幼树在中南部分布较密集(图3-a)。其中,Ⅰ级(0 mm 样地2中幼苗幼树呈斑块分布在整个样地,密度远大于样地1(图3-b)。调查显示,样地2中较多的枯立木和倒木对幼苗幼树的分布存在一定的影响。地径Ⅰ级(0 mm 样地1(图4-c)和样地2(图4-d)幼树幼苗在每个高度级上的个体数差异没有地径显著。样地1中,Ⅰ级(H≤0.33 m)和Ⅱ级(0.33 m 4结论 丝栗栲种群在阴阳两个坡向的密度存在差异,阳坡密度(1 612株/hm2)小于阴坡(2 124株/hm2)。丝栗栲种群4个发育阶段的株数分布在阴阳2个坡向的变化趋势一致,2个样地均为Ⅰ级立木最多,分别占总立木的50.87%、60.45%,依次减少,Ⅳ级立木最少,分别占总立木的6.70%、4.52%。从4个发育阶段来看,样地2幼苗和幼树的株数远大于样地1,但中树和大树的株数略少于样地1。说明阳坡地段幼苗经过自疏和他疏作用进入幼树阶段后,幼树又能以较强的竞争力进去中树阶段,这和阳坡林分密度和较为充足的光照等环境资源有关;而阴坡地段林下荫蔽的环境为丝栗栲幼苗幼树创造了良好的生长条件,但在进入中树和成树阶段时,由于光照等资源的限制,种内竞争加剧,使得种群死亡率远高于阳坡。不同发育阶段活立木比例存在差异,表现为幼树幼苗多,中树大树少,种群直径分布为倒“J”形,属于增长型种群,幼苗贮备量大,天然更新良好。 更新幼苗和幼树的数量随地径和树高的增大呈递减趋势。丝栗栲Ⅰ级幼苗数量最多,Ⅱ级幼苗数量次之,Ⅵ、Ⅶ、Ⅷ 级幼树数量最少,更新幼苗幼树株数随地径级的增加呈降低趋势,地径分布曲线呈倒“J”形,幼树幼苗结构为增长型,对于幼树来说,幼苗的补给充足。样地1的更新苗木高度结构曲线呈明显的左偏山状分布,而样地2呈近似双峰型分布,整体来看,丝栗栲幼苗幼树的树高结构呈明显的非对称性山状曲线,接近幼苗幼树平均高的林木株数较多,Ⅱ、Ⅲ、Ⅳ级占总株数的53.04%,Ⅶ、Ⅷ级株数较少。幼树幼苗在每个高度级上的个体数差异没有地径显著。空间分布上,更新幼苗幼树在阳坡下坡位分布多,在阴坡呈斑块状分布。 参考文献: [1]李霖. 福建省永安市贡川镇低产丝栗栲次生林特征[J]. 山地学报,2007,25(3):333-337. [2]潘尧燕. 低产丝栗栲次生林林分特征研究[J]. 林业勘察设计,2010(1):74-76. [3]顾红梅,邓光华,翁贤权,等. 丝栗栲幼苗对不同氮含量及形态的营养响应[J]. 江西农业大学学报,2016,38(2):312-318. [4]姜汉侨,段昌群,杨树华,等. 植物生态学[M]. 北京:高等教育出版,2004:157-159.[HJ1.77mm] [5]Frost I,Rydin H. Spatial pattern and size distribution of the animall dispersed tree Quercusrobur in two spruce-dominated forests[J]. Ecoscience,2000,7(1):38-44. [6]閆淑君,洪伟,吴承祯,等. 丝栗栲种群生命过程及谱分析[J]. 应用与环境生物学报,2002,8(4):351-355. [7]杨华,李艳丽,沈林,等. 长白山云冷杉林幼苗幼树空间分布格局及其更新特征[J]. 生态学报,2014,34(24):7311-7319. [8]王伯荪,余世孝,彭少麟,等. 植物群落学实验手册[M]. 广州:广东高等教育出版社出版,1996. [9]汪洋,张敏,陈霞,等. 七姊妹山保护区红椿种群动态与谱分析[J]. 湖北农业科学,2015,54(22):5632-5636. [10]樊登星,余新晓. 北京山区栓皮栎林优势种群点格局分析[J]. 生态学报,2016,36(2):318-325. [11]巢林,洪滔,林卓,等. 中亚热带杉阔混交林直径分布研究[J]. 中南林业科技大学学报,2014,34(9):31-37. [12]丁国泉,许继中. 辽东山区天然次生林林分直径分布模型研究[J]. 林业资源管理,2012(5):94-97,124.
猜你喜欢
南水北调与水利科技(2017年1期)2017-02-27中国人口·资源与环境(2016年11期)2017-02-17绿色科技(2016年20期)2016-12-27地球科学与环境学报(2015年4期)2015-07-31
