
混合物料干湿厌氧发酵特性及菌群结构
2019-08-20 14:58郑讯涛王乐乐张寓涵张洋洋潘云霞
江苏农业科学 2019年8期
郑讯涛 王乐乐 张寓涵 张洋洋 潘云霞
摘要:厌氧发酵是解决秸秆和畜禽粪等农业废弃物污染的重要途径,为实现秸秆和牛粪的资源化利用,研究稻秆和牛粪混合物料干、湿厌氧发酵特性,并对混合物料发酵前后的菌群结构进行分析。结果表明,稻秆与牛粪混合物料干、湿发酵的挥发性脂肪酸(VFAs)均以乙酸和丙酸为主,发酵pH值均在5.6~7.7之间变化,湿发酵的总固体浓度(total solid,简称TS)累积产甲烷量(63.8 mL/g)较干发酵(36.9 mL/g)提高73.90%,发酵前后菌群结构发生较大变化,细菌的优势菌群由消化链球菌(Peptostreptococcaceae)、瘤胃菌(Ruminococcaceae)、梭菌(Clostridiaceae.1)和理研菌(Rikenellaceae)轉变为普雷沃氏菌(Prevotellaceae)、毛螺菌(Lachnospiraceae)和互养菌(Synergistaceae),古菌的优势菌群由甲烷杆菌(Methanobacteriaceae)和甲烷球菌(Methanosarcinaceae)转变为以乙酸代谢为主的甲烷球菌(Methanosarcinaceae)。
关键词:稻秆;牛粪;混合物料;厌氧发酵;特性;菌群结构
中图分类号: X71;S188+.4文献标志码: A
文章编号:1002-1302(2019)08-0238-04
我国每年产生的秸秆和畜禽粪达到8.00、32.64亿t[1],如果未能妥善处理,会对生态环境造成巨大危害。以秸秆与畜禽粪混合为底物进行厌氧发酵生产沼气,既可解决农村畜禽粪处理原料不足问题,也可缓解环境污染压力,还能实现清洁能源的生产[2-3]。目前秸秆与畜禽粪的厌氧发酵多采用湿式厌氧发酵形式,该方法耗水量大,沼液产生量多[4],后续处理不便。干发酵由于其发酵过程需水量较少,发酵后无沼液产生、沼渣可用于直接制有机肥、减少二次污染等优点受到普遍关注[5-6];但单纯的畜禽粪干发酵,由于物料氨态氮含量高,发酵系统易产生氨中毒现象[7-8],而且物料较差的透气性和传质性能也使发酵系统效率较低[9]。单一秸秆干发酵仍须添加额外氮源以防止酸化现象[10];将畜禽粪与秸秆混合进行干发酵既可改善发酵体系中的C/N平衡,降低发酵过程中氨抑制的风险,又可通过秸秆的疏松结构改善原料间的接触面、增强物料的传质性能。在发酵中微生物菌群对沼气的生产发挥了重要作用[11-12],分析厌氧发酵过程中微生物菌群的结构及代谢产物的变化有助于了解发酵系统的运行状态,便于改进发酵工艺,提高发酵系统的厌氧消化效率及产气量。为此,本研究将鲜牛粪与稻秆按比例混合,研究混合物料高含固率的湿发酵总固体浓度(total solid,简称TS)(TS=10%)与干发酵(TS=25%)的厌氧消化特性,并对发酵潜力好的一组细菌和古菌微生物菌群进行16S rRNA高通量测序,研究发酵前后微生物菌群的结构及功能变化,以期通过厌氧发酵途径为秸秆和畜禽粪的资源化利用提供可靠技术支持。
1材料与方法
试验于2016年11月至2017年3月在西南大学废弃物资源化利用实验室进行。
1.1试验材料
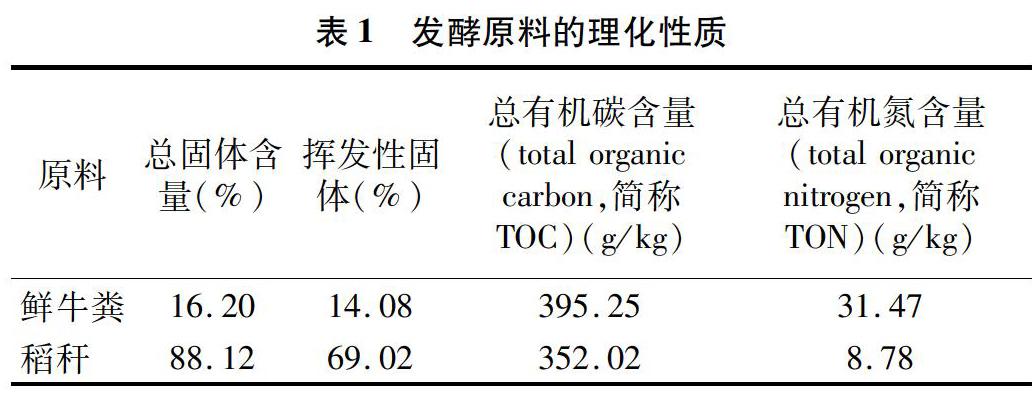
鲜牛粪取自重庆市北碚区某奶牛养殖场;稻秆取自重庆市北碚区歇马镇某稻田,稻秆已在田间自然风干半年,试验前去掉稻穗,并将稻秆剪至1~2 cm的小段。试验材料的理化性质如表1所示。
1.2试验装置与试验方法
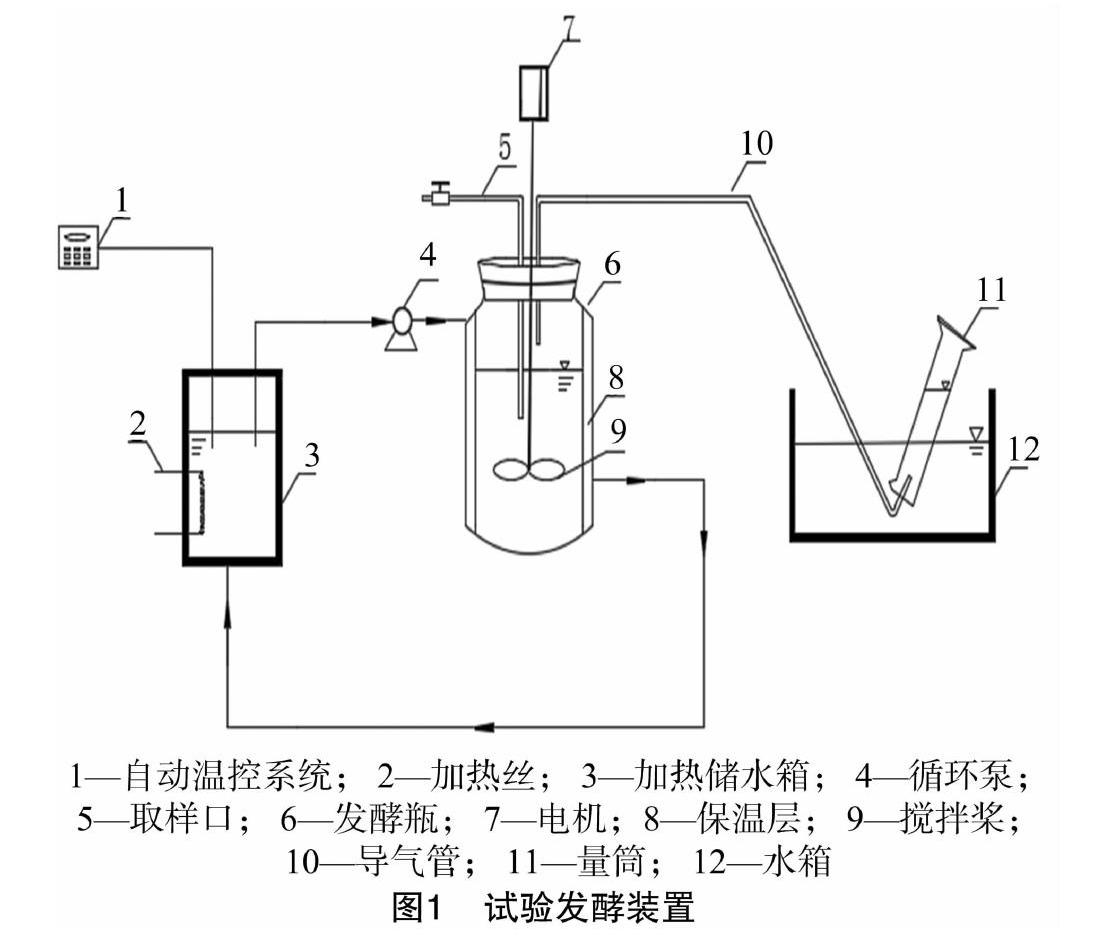
试验采用自制的发酵装置(图1),主要由2.5 L的发酵瓶、加热与温控装置、循环泵、集气量筒、导气管等组成。发酵罐温度通过外部储水箱水加热后泵送至发酵罐保温层,以保持恒温(35±1) ℃,产气量由排水法收集。试验设置干发酵组和湿发酵组,2组试验发酵原料总质量均为800 g,总固体浓度分别为10%、25%,试验中稻秆与鲜牛粪按照干物质比 1 ∶1 添加,厌氧发酵前先通入5 min氮气以除去发酵瓶内的氧气。
1.3分析方法
总固体含量采用烘干法测定;挥发性固体(volatile solid,简称VS)采用马弗炉灼烧法测定[13];pH值采用精密pH计(Sartorius-10型)测定;挥发性脂肪酸(volatile fatty acids,简称VFAs)采用高效液相色谱仪(LC2010)测定;纤维素酶活性采用DNS比色法测定[14];酶活性单位定义:1 mL发酵液在一定pH值和温度下在1 min内催化底物生成1 μmol的葡萄糖为1个酶活单位,用U表示;总产气量采用排水法测定,甲烷含量采用GC9720型气相色谱仪进行分析测定[15]。
1.4微生物多样性的测定
1.4.1总DNA的提取
取摇匀后的1.5 mL发酵液,过滤,12 000 r/min离心5 min,弃上清液,剩余沉淀物用于总DNA提取。总DNA的提取按照试剂盒(OMEGA Soil DNA Kit)的说明书进行,使用0.8%琼脂糖凝胶电泳进行分子大小判断,并利用紫外分光光度计对DNA进行定量分析。
1.4.2微生物菌群结构检测
试验选用长度约为250 bp的细菌16S rRNA基因高度可变的V4区和长度为420 bp的古菌16S rRNA基因高度可变的V4V5区测序。PCR扩增选用细菌16S rDNA V4区特异性引物,520F(5′-barcode+[JP9]AYTGGGYDTAAAGNG-3′)、802R(5′-[JP9]TACNVGGGTATCTAATCC-3′);古菌进行PCR扩增选用的引物为519F(5′-[JP9]CAGCCGCCGCGGTAA-3′)、915R(5′-[JP9]GTGCTCCCCCGCCAATTCCT-3′)。PCR反应体系(25 μL)为:0.25 μL Q5 high-fidelity DNA聚合酶,5 μL 5×Reaction Buffer,5 μL 5×High GC Buffer,2 μL dNTP(10 mmol/L),2 μL DNA模板,1 μL正向引物(10 μmol/L),1 μL 反向引物(10 μmol/L),加水补足至 25 μL。PCR扩增程序:98 ℃预变性30 s;98 ℃变性15 s,50 ℃ 退火30 s;72 ℃延伸30 s,25个循环;72 ℃延伸5 min。将扩增产物通过2%琼脂糖凝胶电泳进行检测,目标片段采用上海百赛生物技术股份有限公司(Axygen)的凝胶回收试剂盒进行回收。文库的制备和高通量测序由测序公司进行。
2结果与分析
2.1干、湿发酵过程中VFAs与pH值的变化
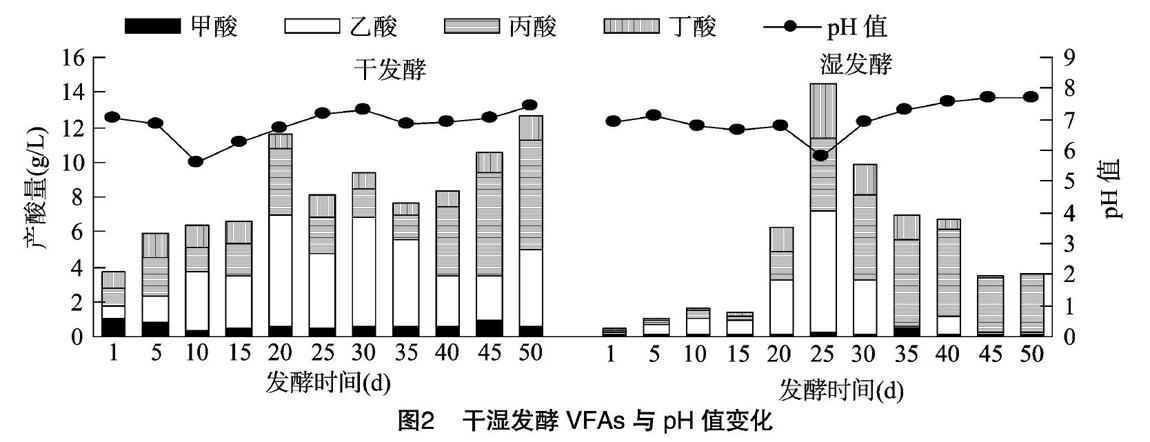
pH值和VFAs对厌氧消化产沼气有重要影响[16-17]。由图2可知,在稻秆与牛粪混合物料的干、湿发酵中,VFAs与pH值的变化存在较大差异。干发酵的pH值在发酵10 d时降至最低,随后处于逐渐升高状态,而VFAs达到峰值的时间滞后于pH值降低的时间,但湿发酵的pH值降低与VFAs的升高同步。在稻杆与牛粪混合物料的干、湿发酵中,总VFAs均以乙酸和丙酸为主,甲酸和丁酸的含量相对较少,且乙酸含量随发酵时间均呈先上升后下降的趋势。在干发酵中,乙酸的平均浓度占总VFAs的44.4%,明显高于湿发酵的30.5%,丙酸的平均浓度为2.85 g/L,高于产甲烷菌对丙酸的耐受浓度1.0 g/L[18],所以在干发酵后期出现了丙酸积累。湿发酵进行到40 d时,丙酸含量累积到5.04 g/L,远超过产甲烷菌的耐受能力,但由于此时甲酸、乙酸、丁酸浓度很低,加之湿发酵较好的传质能力,产甲烷菌能够间接利用丙酸进行代谢[19],使丙酸含量迅速降低,所以虽然发酵结束丙酸含量占总VFAs的90%,但系统并没有出现酸化现象。
2.2干、湿发酵过程中酶活性的变化
混合发酵物料中含有半纤维素、纤维素、木质素,纤维素酶活性在一定程度上反映了发酵底物的降解效率。由图3可知,复合酶系的3种酶均参与混合物料的发酵水解,均呈先升高后降低的趋势,并且在3种酶中酶活性最高的是内切葡聚糖酶与β-葡萄糖苷酶,外切葡聚糖酶虽在发酵中期有较大活性,但低于内切葡聚糖酶和β-葡萄糖苷酶活。干、湿发酵的滤纸总酶活性在发酵的前20 d整体呈上升趋势,并在20 d时达到峰值1.68、1.40 U/TS,随后整体呈下降趋势。
2.3干、湿发酵过程中日产气量与累积产气量的变化
由图4-a可知,混合物料干发酵与湿发酵在最初5 d均经历了1个小的产气高峰(320、415 mL),但此时气体成分中甲烷含量均较低(图4-b)。湿发酵在30、41 d时出现了2次产气高峰,产气量分别达到765、635 mL,干发酵在28 d时产气量开始增加,43 d时达到产气高峰909 mL,之后逐渐下降,发酵结束时,湿发酵累积TS甲烷产量为63.8 mL/g,较干发酵的甲烷产量高73.2%(图4-b)。为此,对湿发酵的微生物菌群结构进行分析。
2.4混合物料湿发酵中微生物菌群分布
由图5可知,厌氧发酵前后,细菌和古菌在菌群丰度上发生了较大的变化。在发酵初期(发酵5 d),细菌中消化链球菌(Peptostreptococcaceae)、瘤胃菌(Ruminococcaceae)、梭菌(Clostridiaceae.1)、理研菌(Rikenellaceae)是优势菌群,分别占27.6%、15.8%、12.6%、6.7%;经厌氧消化后,普雷沃氏菌(Prevotellaceae)、克里斯腾森菌(Christensenellaceae)、毛螺菌(Lachnospiraceae)、互养菌(Synergistaceae)较发酵初期分别提高16.4%、13%、7%、5.8%,成为产气高峰(发酵40 d)的优势细菌菌群。古菌中,发酵初期(发酵5 d)的产甲烷菌主要有甲烷杆菌(Methanobacteriaceae)与甲烷球菌(Methanosarcinaceae),它们的丰度差别不大,分别占菌群总数的 16.2%、17.5%,但发酵进行至产气高峰(发酵40 d)后,甲烷球菌成为主要的优势菌群,其丰度达到33.8%,而甲烷杆菌降至9.2%。
3讨论
糖类是厌氧发酵过程中的重要中间产物,是多数微生物的营养物质。在发酵底物主要为木质纤维素时,糖类的产生主[CM(25]要依赖于微生物分泌的纤维素酶对木质纤维素的水解作用。试验中,发酵前期纤维素酶活性逐渐升高,通过菌群检测发现能够分泌纤维素酶的瘤胃菌(Ruminococcaceae)[20-21]、能够利用木聚糖和淀粉等物质的普雷沃氏菌(Prevotellaceae)[22]、克里斯腾森菌(Christensenellaceae)、毛螺菌(Lachnospiraceae)及互养菌(Synergistaceae)成为优势菌群,同时,能够通过糖酵解平台水解产生VFAs的梭菌(Clostridiaceae.1)和理研菌(Rikenellaceae)也是优势菌群[23-24],因此发酵前期VFAs的含量随纤维素酶活性升高而增加。在发酵中后期,利用H2/CO2、甲酸盐、2-丙醇/CO2、2- 丁醇/CO2的甲烷杆菌[25]丰度大幅下降,利用乙酸和甲醇等營养物质[26]的甲烷球菌成为优势菌群,VFAs中的乙酸被甲烷球菌利用产生甲烷,所以此时发酵系统中,乙酸含量几乎为0,而甲烷含量逐渐增加。研究表明,只有当发酵系统内的氢分压稳定在 0.1~10.1 Pa范围内时,丙酸的降解才会发生[27],但发酵后期氢养型甲烷短杆菌的丰度大幅下降,不能维持系统内较低的氢分压,这可能是丙酸积累的重要原因。
4结论
稻秆与牛粪混合物料在干、湿发酵过程中,VFAs均以乙酸和丙酸为主,甲酸和丁酸含量相对较低,pH值为5.6~7.7,发酵系统没有出现酸化现象,湿发酵的累积TS甲烷产量(63.8 mL/g)较干发酵(36.9 mL/g)提高72.90%。稻秆与牛粪混合物料的湿发酵以甲烷球菌(Methanosarcinaceae)的乙酸代谢为主,发酵前后,微生物菌群结构发生较大变化。发酵初期,细菌菌群以消化链球菌(Peptostreptococcaceae)、瘤胃菌(Ruminococcaceae)、梭菌(Clostridiaceae.1)和理研菌(Rikenellaceae)为主,甲烷杆菌(Methanobacteriaceae)与甲烷球菌(Methanosarcinaceae)为古菌优势菌群,厌氧发酵后,细菌优势菌群转变为普雷沃氏菌(Prevotellaceae)、毛螺菌(Lachnospiraceae)和互养菌(Synergistaceae),古菌则以甲烷球菌(Methanosarcinaceae)为主。
参考文献:
[1]贾伟. 我国粪肥养分资源现状及其合理利用分析[D]. 北京:中国农业大学,2014:1-19.
[2]魏泉源. 规模化沼气工程沼液、沼渣减量化及资源化利用研究[D]. 北京:北京化工大学,2014:1-12.
[3]宋香育,张克强,房芳,等. 工艺措施对猪粪秸秆混合厌氧干发酵产气性能的影响[J]. 农业工程学报,2017,33(11):233-239.
[4]马旭光,李传友,袁旭峰,等. 高含固率秸秆和牛粪混合物料发酵产甲烷工艺[J]. 农业工程学报,2014,30(14):227-235.
[5]Li Y B,Park S Y,Zhu J Y. Solid-state anaerobic digestion for methane production from organic waste[J]. Renewable & Sustainable Energy Reviews,2011,15(1):821-826.
[6]Abbassi-Guendouz A,Brockmann D,Trably E,et al. Total solids content drives high solid anaerobic digestion via mass transfer limitation[J]. Bioresource Technology,2012,11(3):55-61.
[7]馮磊,李润东. 牛粪单级干发酵产气中试研究[J]. 农业环境科学学报,2011,30(11):2374-2378.
[8]盛迎雪,曹秀芹,张达飞,等. 猪粪干式厌氧消化中试试验研究[J]. 中国沼气,2016,34(5):41-46.
[9]李文哲,罗立娜,窦钰程,等. 干湿耦合厌氧发酵工艺设计与发酵特性试验[J]. 农业机械学报,2013,30(5):148-153.
[10]方玉美,任秋鹤,聂宁,等. 秸秆高温干式厌氧发酵的产气特性研究[J]. 河南科学,2017,35(6):891-896.
[11]于美玲,于洋,董晓莹,等. 城市有机垃圾厌氧干发酵菌群与代谢产物特性研究[J]. 中国沼气,2015,33(6):14-18.
[12]虞方伯,罗锡平,管莉菠,等. 沼气发酵微生物研究进展[J]. 安徽农业科学,2008,36(35):15658-15660.
[13]刘战广,朱洪光,王彪,等. 粪草比对干式厌氧发酵产沼气效果的影响[J]. 农业工程学报,2009,25(4):196-200.
[14]鲍文英,江经纬,周云,等. 一株木质纤维素降解菌的筛选及其全基因组分析[J]. 微生物学报,2016,56(5):765-777.
[15]Li D,Liu S,Mi L,et al. Effects of feedstock ratio and organic loading rate on the anaerobic mesophilic co-digestion of rice straw and pig manure[J]. Bioresource Technology,2015,187:120-127.
[16]Lim S J,Kim B J,Jeong C M,et al. Anaerobic organic acid production of food waste in once-a-day feeding and drawing-off bioreactor[J]. Bioresource Technology,2008,99(16):7866-7874.
[17]Cavinato C,da Ros C,Pavan P,et al. Influence of temperature and hydraulic retention on the production of volatile fatty acids during anaerobic fermentation of cow manure and maize silage[J]. Bioresource Technology,2017,223:59-64.
[18]王艳芹,付龙云,杨光,等. 农村有机生活垃圾等混合物料厌氧发酵产沼气性能[J]. 农业资源与环境学报,2016,35(6):1173-1179.
[19]凡慧,马诗淳,黄艳,等. 丙酸互营氧化菌群对厌氧消化过程中丙酸累积的调控研究进展[J]. 中国沼气,2016,34(4):3-7.
[20]Wood T M,Wilson C A,Stewart C S. Preparation of the cellulase from the cellulolytic anaerobic rumen bacterium,Ruminococcus albus,and its release from the bacterial cell wall[J]. Biochemical Journal,1982,205(1):129-137.
[21]Doerner K C,White B A. Assessment of the endo-1,4-beta-glucanase components of Ruminococcus flavefaciens FD-1[J]. Applied & Environmental Microbiology,1990,56(6):1844-50.
[22]Purushe J,Fouts D E,Morrison M,et al. Comparative genome analysis of Prevotella ruminicola and Prevotella bryantii:insights into their environmental niche[J]. Microbial Ecology,2010,60(4):721-729.
[23]Zened A,Combes S,Cauquil L,et al. Microbial ecology of the rumen evaluated by 454 GS FLX pyrosequencing is affected by starch and oil supplementation of diets[J]. FEMS Microbiology Ecology,2013,83(2):504-514.
[24]Islam R,Cicek N,Sparling R,et al. Effect of substrate loading on hydrogen production during anaerobic fermentation by Clostridium thermocellum 27405[J]. Applied Microbiology & Biotechnology,2006,72(3):576-583.
[25]方晓瑜,李家宝,芮俊鹏,等. 产甲烷生化代谢途径研究进展[J]. 应用与环境生物学报,2015,21(1):1-9.
[26]承磊,郑珍珍,王聪,等. 产甲烷古菌研究进展[J]. 微生物学通报,2016,43(5):1143-1164.
[27]赵杰红,张波,蔡伟民. 厌氧消化系统中丙酸积累及控制研究进展[J]. 中国给水排水,2005,21(3):25-27.
猜你喜欢
意林彩版(2022年2期)2022-05-03
煤气与热力(2021年12期)2022-01-19
今日农业(2021年10期)2021-11-27
趣味(语文)(2020年11期)2020-06-09
贵州大学学报(自然科学版)(2019年5期)2019-10-23
小哥白尼(野生动物)(2019年6期)2019-10-10
中国资源综合利用(2018年12期)2019-01-08
农业工程学报(2017年21期)2017-11-20
广东农业科学(2017年5期)2017-08-29
浙江农业科学(2016年11期)2016-05-04
