
沙漠腹地达理雅博依天然绿洲植物群落特征分析∗
2019-08-27 11:25张文奇周小龙尼加提卡斯木迪丽尼格尔艾合买提比拉力依明李浩师庆东
新疆大学学报(自然科学版)(中英文) 2019年3期
张文奇,周小龙,尼加提·卡斯木,迪丽尼格尔·艾合买提,比拉力·依明,李浩,师庆东
(1.新疆大学资源与环境科学学院,新疆乌鲁木齐830046;2.新疆大学干旱生态环境研究所,新疆乌鲁木齐830046;3.绿洲生态教育部重点实验室,新疆乌鲁木齐830046)
0 引言
植物群落是同一自然区域内不同植物经过长时间的相互作用、相互适应和环境筛选后形成的集合,是生态系统的主要组成部分,它为人类、动植物及其他生物提供了栖息场所和食物来源[1].一个地区植物群落的整体状况综合体现了该地区的生态本底,是进行生态恢复、物种多样性资源保护和制定土地利用政策的重要依据[2].在生态系统较为脆弱的情况下,许多植物物种很容易因某一环境因子的改变而衰退或消亡,因此植物群落的物种组成、分布格局、种群结构的变化可以对生态环境的演变趋势起到指示作用[3].
达理雅博依绿洲位于新疆维吾尔自治区于田县境内[4],主要包括克里雅河下游河道及天然尾闾绿洲,不同于其他环绕式分布在沙漠边缘的绿洲,其是世界上罕见的沙漠腹地规模化天然绿洲[5,6].深入塔克拉玛干沙漠腹地的达理雅博依绿洲是我国极端干旱区沙漠绿洲生态系统的典型代表,生态系统极其脆弱,但生态功能极为重要,对塔克拉玛干南缘绿洲环境的维护具有不可替代的作用[7].
长期以来由于自然条件恶劣、位置偏远和交通不便等原因,关于该绿洲植物群落特征的系统研究很少.本研究通过实地样方调查的方式对该区域植物群落物种组成、常见功能性状特征、种群年龄结构、植被健康状况和种间关系进行了分析和讨论.本文可以为该绿洲后续的相关研究提供有价值的群落特征基础信息,同时,在提倡“绿水青山就金山银山”[8]的时代大背景下,为绿洲日后的生态经济发展规划、生态环境及生态系统功能的维持提供可靠的科学依据.
1 研究区概况
新疆于田县达里雅博依绿洲地处世界第二大流动沙漠——塔克拉玛干沙漠腹地,是由克里雅河下游形成的叶状尾闾绿洲[9].该地区纵深塔克拉玛干大沙漠腹地 250 km,地理坐标为 81◦41′15′′∼82◦9′41′′E,38◦14′39′′∼ 38◦37′35′′N,地势南高北低,地形呈南北长带状(图1).绿洲长80 km、宽7∼15 km,面积约为342 km2,海拔高度1 161∼1 212 m[10].
该区为暖温带干旱沙漠气候,蒸发量大,降雨少,光热资源充足,全年太阳总辐射量为143 Col·cm−2,全年≥10℃的积温4 208℃,无霜期192∼235 d.研究区气温年际间变化比较稳定,年平均气温11.6℃,气温的年较差和日较差较大.降水不足20 mm,但年平均蒸发量为2 480 mm,为降水的124倍,大气极度干燥.研究区浮尘、沙暴天气较多,平均大风日2.1 d,年均沙暴日数20.3 d,年均浮尘日数155.4 d[11].克里雅河下游沿岸的植被主要为胡杨(Populus euphratica),柽柳(Tamarix)以及芦苇(Phragmites communis)等[12],这些植被形成了一道屏障,对荒漠的扩张起到了阻碍作用.

图1 研究区示意图Fig 1 Research area location
2 研究方法
2.1 野外样方调查
本研究采用样方调查的方式获取植被信息,样方位置预先由卫星地图定点,保证样点在整个绿洲大致均匀分布,共计26个(图2).样方调查于2018年7月中旬进行,在事先定好的样点周围随机选取一个20 m×20 m的样方,记录样方内出现的所有乔木和灌木的个体数量,然后在该样方内嵌套一个1 m×1 m的小样方,对其中草本植物的多度和盖度进行调查.同时我们随机选取植物个体(如果个体数量少于15个则全部调查)进行功能性状的测定,乔木的主要指标为胸径、株高、冠幅和枯枝率;灌木的主要指标为株高、冠幅和基径;草本测量的指标为高度.调查过程中详细记录各样点的编号,所有样方采用GPS定位,拍照存档建立资料库,采集植物标本存档并结合有关达理雅博依地区的植物文献和植物志进行鉴定和分析[13].

图2 绿洲样方示意图Fig 2 Locations of quadrats in the Daliyaboyi Oasis
2.2 植被参数特征
为了客观描述达理雅博依绿洲典型植被类型(胡杨、柽柳和芦苇)空间格局的差异性,本文根据绿洲的自然形状、植被和水文条件将绿洲分为三个部分(北部、中部和南部),并基于野外实地植被调查数据,采用以下指数对绿洲植被参数进行特征分析.(由于绿洲内柽柳种类复杂,为了更好地进行计算和比较,按照柽柳属进行统计分析)
相对频度=某个种的频度/所有种的频度之和×100%
相对多度=某个种的多度/所有种的多度之和×100%
相对盖度=某个种的盖度/所有种的盖度之和×100%
相对高度=某个种的平均高度/所有种的平均高度之和×100%
2.3 种群结构和健康程度分析方法
基于以上研究,本文采取立木结构代表树木年龄的方法[14],依据树木胸径(DBH,diameter at breast height)大小依次将胡杨种群划分为Ⅰ(DBH≤4 cm)、Ⅱ(4 cm<DBH<10 cm)、Ⅲ(10 cm≤DBH<20 cm)、Ⅳ(20 cm≤DBH<30 cm)、Ⅴ(30 cm≤DBH≤50 cm)、Ⅵ(DBH>50 cm)共6个等级,对应林龄为幼苗(Ⅰ)、幼龄林(Ⅱ)、中龄林(Ⅲ-Ⅳ)、成熟林(Ⅴ)和过熟林(Ⅵ).按树木高度将胡杨种群分为Ⅰ(H<5 m)、Ⅱ(5 m≤H<10 m)、Ⅲ(10 m≤H<15 m)、Ⅳ(H≥15 m)共4个等级.
通常一个地区植被的健康状况可以表征当地环境的优劣程度.为了获取胡杨的健康状况,根据植被样方调查得到的胡杨枯枝率,将胡杨健康程度划分为死亡V5(立木无活枝或倒下)、濒死V4(仅有部分活枝)、衰退V3(枯枝占比达50%以上)、中等V2(有部分枯枝)、健康V1(无枯枝或有少量枯枝)5个等级(表1).

表1 胡杨(Populus euphratica)树木活力的分类标准Tab 1 Classification criteria for tree vigor of Populus euphratica
柽柳植株生长缓慢,其基径与树龄成正比,因此我们研究柽柳基径(BD,basal diameter)与株高这两个种群性状的结构特征与空间分布规律[15].分级统计柽柳株数,以株数为纵坐标,基径大小为横坐标,根据柽柳生活史特征,以1 cm为分级标准,BD≤1 cm为A级,1 cm<BD<2 cm为B级,C级(2 cm≤BD<3 cm),以此类推,BD≥14 cm为O级,共15级.对不同径级柽柳频数取自然对数,进而绘制柽柳存活曲线.
2.4 种间关系分析
种间关联度采用2×2列联表法计算关联系数(Dagnelie系数)[16],表中a表示兼有物种A、B的样方数量,b表示有B无A的样方数量,c表示有A无B的样方数量,d表示两者均无的样方数量(表2).

表2 种间关联度计算Tab 2 Inter-species correlation calculation
Dagnelie系数可以表示为:

上式中,关联系数V的取值范围为[-1,1],一般认为:V>0时,表明物种A与物种B正关联;V≤0时,表明物种A与物种B负关联.

公式(1)与公式(2)中的字母含义与表2中相同.
3 结果与分析
3.1 植物群落物种组成与系统发育信息
根据此次样方调查结果对达理雅博依绿洲内的植物进行分类统计,结果表明,该绿洲内的植物均为被子植物,其中双子叶植物有9科,单子叶植物只有1科(表3).该绿洲内的植物物种数共计8目10科15属18种.

表3 达理雅博依绿洲植物整体统计Tab 3 Whole plants statistic on the Daliyaboyi Oasis
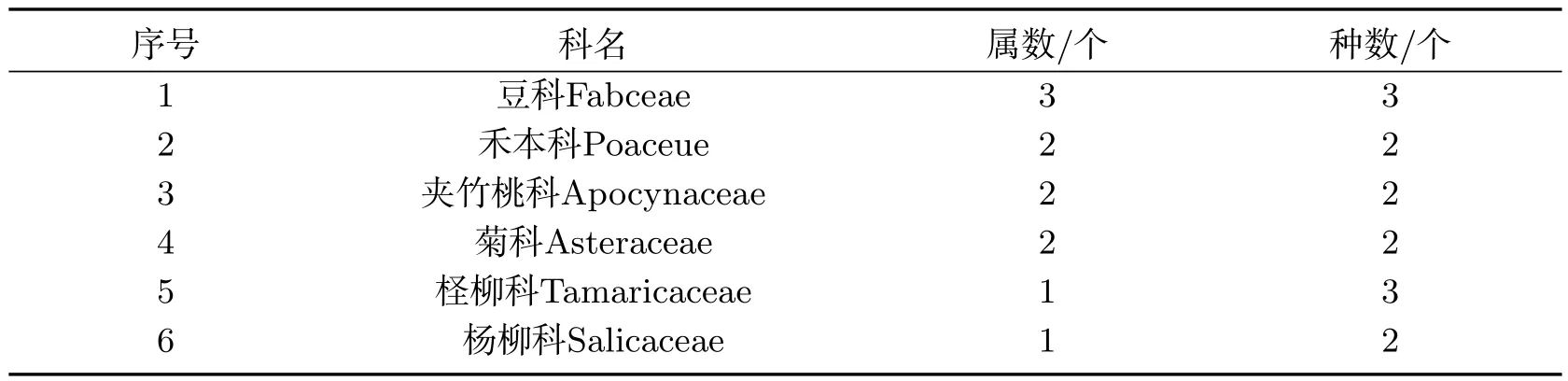
表4 达理雅博依绿洲主要优势科Tab 4 Main advantage families of the Daliyaboyi Oasis

表5 达理雅博依绿洲科、属、种统计Tab 5 Family,genus and species statistic on the Daliyaboyi Oasis
根据表4和表5的统计可知,达理雅博依绿洲植物构成中,柽柳科和豆科占全绿洲总科数的20.00%、总属数的26.67%、总种数的33.33%(两科物种数最多,均为3种).绿洲内的优势科(含物种数2种)有4个,分别是夹竹桃科、禾本科、杨柳科和菊科,占总科数的40.00%,总属数的46.67%,总种数的44.44%.单科单属种的共4科4属4种,分别占整个绿洲植物总科数、总属数、总种数的百分比为40.00%、26.67%和22.22%.
达理雅博依绿洲内植物物种组成较为单一,样方调查所得整个绿洲植物种类共有18种(表6),主要优势种为胡杨、柽柳和芦苇.总体而言,本绿洲群落结构简单,以芦苇、花花柴这类多年生草本为建群种的小群落一般只有一个层片,没有其他的伴生种.以柽柳为建群种的灌木群落的层片只有1∼2个,群落结构也较简单.以胡杨为优势种的群落包含乔木层、灌木层和多年生草本层,但有些地方随着地下水位的下降,林下植被逐渐消失,形成了仅剩胡杨的单一乔木层,并且这种单一的群落结构在绿洲样方调查中时常见到.

表6 达理雅博依绿洲内植物种统计Tab 6 Species statistic on the Daliyaboyi Oasis
3.2 绿洲内部群落优势种的多度、盖度和高度的变化规律
本文按照绿洲内的植被状况和环境变化梯度将绿洲划分为北部、中部和南部三个部分,并分别对这三个部分的植物群落组成和功能性状特征进行分析比较.结果显示,从植被基本参数的南北方向变化趋势来看,胡杨的相对多度、相对密度和相对高度等指数的均值由南向北都呈上升趋势,尤其是相对高度的变化趋势最为明显.无论从变化幅度,还是均值的角度来考虑,胡杨的相对盖度占比都较高,均大于40%,其中北部相对盖度占比最高(大于90%),是绿洲北部植被覆盖度的主要贡献者(图3).
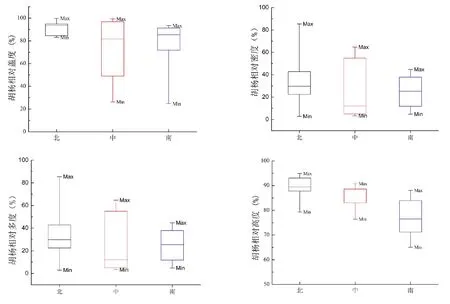
图3 胡杨植物参数特征Fig 3 Plant parameters of Populus euphratica
柽柳相对多度和相对密度的均值,中部和南部明显高于北部,在中部达到最大值;相对盖度由南向北呈下降趋势,而相对高度在南部最高、北部最低(图4).

图4 柽柳植物参数特征Fig 4 Plant parameters of Tamarix
作为绿洲典型的多年生草本群落,芦苇主要分布在绿洲南部,其相对多度、相对密度和相对高度的均值分别为9.65%、9.60%和28.66%(图5).

图5 芦苇植物参数特征Fig 5 Plant parameters of Phragmites communis
3.3 群落优势种种群年龄结构和健康状况分析
3.3.1 胡杨种群年龄结构分析
达理雅博依绿洲胡杨种群结构为倒金字塔形,幼苗数量较少,成熟个体数量多,属于衰退型种群.Ⅰ级幼苗占比为5.79%,Ⅱ级幼龄林占比为7.89%,Ⅲ-Ⅳ级中龄林占比为31.60%,Ⅴ级成熟林占比为22.10%,Ⅵ级过熟林占比高达32.63%.从树高来看,胡杨平均高度为7.9 m,最高达17.4 m;Ⅰ级胡杨占比为17.80%,Ⅱ级胡杨占比达55.00%,Ⅲ级胡杨占比为25.00%;高度在15 m以上的胡杨占比为2.10%,5∼15 m高度范围的胡杨占比为80.00%(图6).胡杨种群以成熟林为主,幼林占比较少,也反映出胡杨种群更新困难的现状.

图6 胡杨径级与高度结构Fig 6 Diameter class and height structure of Populus euphratica
由胡杨健康分级频数(图7)分析可得,绿洲调查样方内,衰退型胡杨数量占绿洲内胡杨总数的比例最高(40.00%),倒下死亡的胡杨以及枯立木数量占比为29.00%.通体绿叶且几乎没有枯枝的健康型胡杨的数量很少,这说明达理雅博依绿洲现在的植被生存环境整体恶化,如不及时采取措施进行有效地保护与治理,绿洲内部的胡杨将会出现大面积的死亡.

图7 胡杨健康分级频数图Fig 7 Health rating frequency of Populus euphratica
3.3.2 柽柳种群年龄结构分析
柽柳径级-频数关系如图8 a所示,柽柳种群龄级分布为幼龄个体多、中老龄个体少,分布形态为金字塔,反映出柽柳种群具有高繁殖率,这一时期种群处于增长上升阶段.由存活曲线(图8 b)可知,随着径级(龄级)的增加,生存曲线总体呈逐渐下降的趋势,E点和F点呈显著下降趋势,这种现象可能与当地独特的生境息息相关.

图8 柽柳径级与存活曲线Fig 8 Diameter and survival curve of Tamarix
3.4 胡杨与柽柳频度之间存在显著正相关关系
根据野外植被调查样方数据,结合上述关联度计算方法得表7.由公式(1)、(2)计算得到V=0.699,X2=6.84,表示柽柳与胡杨的种间关系为极显著正相关.这种正相关关系暗示两个物种之间可能存在相互促进作用.

表7 胡杨(Populus euphratica)与柽柳(Tamarix)关联度分析Tab 7 Analysis of the correlation between Populus euphratica and Tamarix
4 讨论与结论
4.1 讨论
达理雅博依绿洲植物区系组成简单,这些科的属和种都很少,在潘晓玲对塔里木盆地植物区系的研究中提到“尽管这些科是世界广布的大科,但在本地区种类却极为匮乏”[17],显然达理雅博依绿洲符合这一特点.荒漠植物经向、纬向的分布格局反映了地理位置从古至今的变化,同时是沙漠植被恢复和保护的重要保证[18].克里雅河从南部开始汇入绿洲,丰富的地表水和较高的地下水位使得较多植物种类在这里出现[19],但到了绿洲北端随着河水流量逐渐减少,草本基本已经消失,只有成熟的胡杨林和柽柳靠着发达的根系才能勉强生存下来,形成的层片结构也多为单一种的乔木或灌木层.
对比来看,达理雅博依绿洲胡杨比其他区域胡杨林龄老且整体健康程度差.韩路等人研究塔里木河中游胡杨径级结构时,将DBH大于22.5 cm的胡杨划分为大树[20];刘普幸等人研究敦煌绿洲胡杨时,将DBH在24 cm以上的划分为老树[21];王金山研究塔里木河下游胡杨时,将DBH小于36 cm的胡杨划分为小树,将DBH在48 cm以上的胡杨作为过熟林或衰老林[22].相较而言,达理雅博依绿洲胡杨老树偏多,整体树龄偏大,反映出该地区胡杨更新速度缓慢.造成这种现状的主要原因是胡杨种子萌发对外界条件要求较为苛刻,据估算单株胡杨的结种量最多可达上亿颗,但是萌发率较低,胡杨种子萌发集中在夏季洪水期,散落的种子如果没有着陆在合适的萌发地,自然光照下会快速失去活性[23],湿润河漫滩上少量萌发的幼苗由于后期缺少地表水的补给也容易枯死.胡杨的另一种更新方式是以根孽分苗的方式进行,适宜生境下,胡杨根部生长出不定芽,其伸出地面形成幼株,根孽苗生长所需水分来源于母株的浅层根系[24],这种方式也受限于地表水可达的特定生境.所以,胡杨幼苗生长对地表水的高度依赖,决定了其分布范围狭小,个体数量少.究其根本,外界环境的恶化是造成胡杨种群的壶形分布的根本原因,主要表现为地表水过境频次减少、漫溢范围缩小及地下水位降低,由此导致幼林难以更新.
胡杨的衰退以及柽柳的增长体现了达理雅博依绿洲植被演替的一个现状.该绿洲除了南部地区有其他类型的半灌木或多年生草本出现,整个绿洲基本由柽柳和胡杨组成.导致胡杨与柽柳之间呈正相关关系的原因如下:1)生境的高度重叠.水分是当地最重要的限制因子,因此在距离河道较近或者洪水可以到达的区域,由于水分状况较好,既适合胡杨生长又适合柽柳生长;2)二者之间存在相互促进作用,成年胡杨主要利用深层地下水而柽柳主要利用浅层地下水,在长期适应沙漠特殊生境的过程中,二者可能在水分和养分的利用方式上存在生态位的分化,进而降低了对资源的竞争.而胡杨根系可以将水分从深层提拉到浅层,这种效应有助于柽柳的存活与生长,需进行更多精细的实验来研究胡杨与柽柳之间的共存机制.
4.2 结论
(1)达理雅博依绿洲植物共有10科15属18种,均为被子植物.
(2)胡杨的相对盖度在整个绿洲范围内占比较高,均大于40.00%,为该绿洲的优势种;从地理位置进行分析,随着绿洲由南向北环境梯度的变化,胡杨的相对多度、相对高度和相对密度均呈逐渐增加的趋势.对柽柳而言,其相对多度和相对密度在绿洲南部和中部明显高于北部,在中部达到最大;柽柳的相对盖度由南向北呈下降趋势,而相对高度在南部最大、北部最小.芦苇主要分布在绿洲南部,其相对多度、相对密度和相对高度的均值分别为9.65%、9.60%和28.66%.
(3)柽柳种群年龄结构表现为金字塔形,幼龄个体多、中老龄个体少,处在一个高繁殖率的时期.胡杨种群年龄结构表现为倒金字塔形,幼苗个体较少,成熟个体多,属于衰退型种群.根据枯枝率来定义胡杨的健康程度,发现衰退型胡杨数量占绿洲内胡杨总数的比例最高(40.00%),死亡的胡杨及枯立木数量占比29.00%,健康型胡杨的数量很少.
(4)利用2×2列联表法计算关联系数(Dagnelie系数),由公式(1)、(2)计算得到V=0.699,X2=6.84,表示柽柳与胡杨的种间关系为极显著正相关.
猜你喜欢
林业勘查设计(2022年1期)2022-02-15
乡村科技(2021年17期)2021-10-20
安顺学院学报(2021年4期)2021-09-16
黑龙江工程学院学报(2020年5期)2020-10-21
河北林业科技(2020年3期)2020-03-23
山东林业科技(2019年6期)2020-01-02
水土保持研究(2019年6期)2019-10-19
植物资源与环境学报(2019年1期)2019-04-02
绿色科技(2017年15期)2017-09-01
中国民族医药杂志(2016年8期)2016-05-09
