
降解脐橙囊衣专用霉的诱变育种及产酶工艺优化研究
2019-09-05 12:12龚灵茜
新农业 2019年16期
龚灵茜
(湖南省长沙市雅礼中学,湖南 长沙 410004)
1 研究背景
笔者从湖南邵阳当地的脐橙主产区的腐烂脐橙、土壤中寻找产果胶酶的菌源。通过紫外诱变技术提高酶的产量,优化诱变菌株产酶培养基及其发酵条件,初步确定酶制剂脱囊衣工艺条件,为果胶酶大规模发酵提供参考依据,并且为生物酶法脱囊衣加工脐橙打下基础。
2 结果与讨论
2.1 菌株的分离筛选
2.1.1 初筛各菌株的菌落形态特征。经连续驯化富集实验后,以富含果胶质的脐橙囊衣果渣及柑橘果渣为唯一碳源及能源的培养基平板,对稀释菌悬液进行培养,初筛结果及菌落形态特征描述见表1。
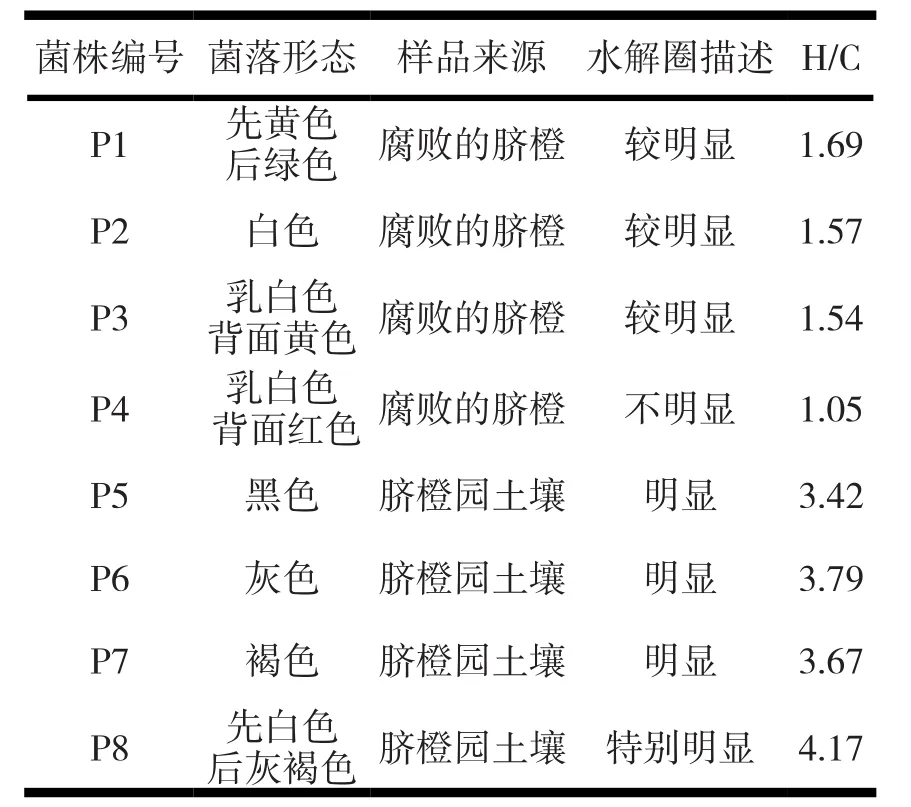
表1 初筛后各菌株的菌落形态
2.1.2 菌株复筛。将透明圈较大的P5、P6、P7、P8菌株,采用复筛培养基进行复筛,进行透明圈H(cm)、生长圈C(cm)值的测定和摇瓶产酶酶活力的定量检测,测定结果见表2。

表2 复筛菌株H/C值与酶活力的比较
由表2复筛测定结果表明,菌株编号为P8酶活力较好;选择该菌株进行分类鉴定。
2.1.3 菌株P8的分类鉴定。菌体形态特征。P8在PDA菌落初为白色,后其上密生一层黑色粉粒,似粗地毯状。菌丝枝繁茂,幼时无隔,老龄有隔;小孢子囊常生在轮生孢囊梗的顶端,孢囊孢子大、色淡、单细胞。
分子生物学鉴定。真菌核糖体基因转录间隔区,又叫内转录间隔区,主要包括内转录间隔区1(ITS1)、5.8S rDNA、内转录间隔区2(ITS2),其两侧分别是18S RNA基因和28S RNA基因。rDNA在种内由于基因的流动而经常表现出很高的同源性,在种间则保持着各种程度的变异。变异的多少能够反映生物进化上属内种间亲缘关系的远近。将菌株P8的rRNAITS2基因序列与GenBank内登录的不同种属霉菌的相应序列进行比对,分析各株序列间的同源性和差异及进化关系。通过上述菌落培养特性、菌体形态观察及18S RNA序列分析,初步鉴定P8为卷枝毛霉。
2.2 菌株P8的诱变及诱变株的筛选
2.2.1 紫外诱变时间。P8孢子紫外诱变剂量效应曲线如图1所示。可以看出,P8孢子经紫外线照射后,再生菌落数逐渐减少。在0~10 min照射时间内,曲线的变化很明显。20 min以后,曲线的下降趋于平缓。根据图1计算出P8最佳诱变的照射剂量为8 min。

图1 紫外诱变剂量效应曲线
2.2.2 诱变菌株的初筛
以8 min作为照射剂量诱变P8孢子,分别诱变3批,每批诱变后的单孢子悬液梯度稀释后涂10个平皿。待孢子在平皿上已再生出菌落后,用灭菌牙签将菌落挑入初筛平板中,每个平板10株诱变菌株,培养2~3 d。为此,从300多株诱变菌株中总共选出40株生长速度快的诱变菌株。
2.2.3 诱变菌株的复筛
分别将40株诱变菌株每株接2个果胶酶筛选斜面培养,计算液体发酵产酶活力。根据酶活力的计算结果,从40株诱变菌株中选出酶活力较高的5株,进行下一步的研究。根据诱变时的原始批次和编号,这5株菌分别命名为Y-9、Y-14、Y-19、Y-26、Y-37。
2.2.4 高产诱变菌株
取上述5株诱变菌株连同原始菌株在接种量相同条件下,进行液体摇瓶发酵产酶培养实验,平行实验3次,产酶结果取平均值,结果见表3。根据表3的结果,选产酶活力明显优于原始菌株的Y-9、Y-26、Y-37编号的3株诱变菌株为高产诱变菌株。

表3 3株诱变菌株及原始菌株液态发酵产酶活力比较

表4 Y-9、Y-26和Y-37以及原始菌株P8连续进行10代发酵产酶培养实验
2.2.5 诱变菌株产酶的遗传稳定
鉴于诱变菌株的性状常不稳定,对得到的3株诱变菌株(Y-9、Y-26和Y-37)进行产酶的遗传稳定性实验,结果见表4。
由表4可看出,Y-9和Y-37分别在传代到第4代、第5代后,产量开始减少,遗传不稳定。Y-26在连续转接的10代里菌株的产酶活力都很稳定,并且都显著超过了对照的原始菌株,其差异均达极显著水平(p<0.01),显示Y-26的遗传性能稳定,可用于后续实验研究。
2.3 诱变菌株Y-26的液体发酵产酶条件
2.3.1 不同碳源与诱变株Y-26的产酶活力。以常见的果胶含量较为丰富的果胶废水、脐橙囊衣果渣、橙皮粉、柚皮粉、桔皮粉为材料。
添加纯果胶及含果胶丰富的果胶废水明显促进诱变株Y-26聚半乳糖醛酸酶、果胶酯酶的合成,表明这两种酶都属于诱导酶。虽然纯果胶作为碳源产酶活力高,但纯果胶价格昂贵,造成生产成本高;橙皮粉、桔皮粉、脐橙囊衣果渣作为碳源产酶更经济,取材范围广,故用脐橙加工废料作为Y-26的碳源来产酶,一方面可以节约成本,另外可避免因加工过程中囊衣及果皮丢弃造成环境污染。
2.3.2 不同氮源与诱变株Y-26产酶活力。本文以硫酸铵等为氮源探讨了诱变株Y-26的产酶活力,结果见表5。
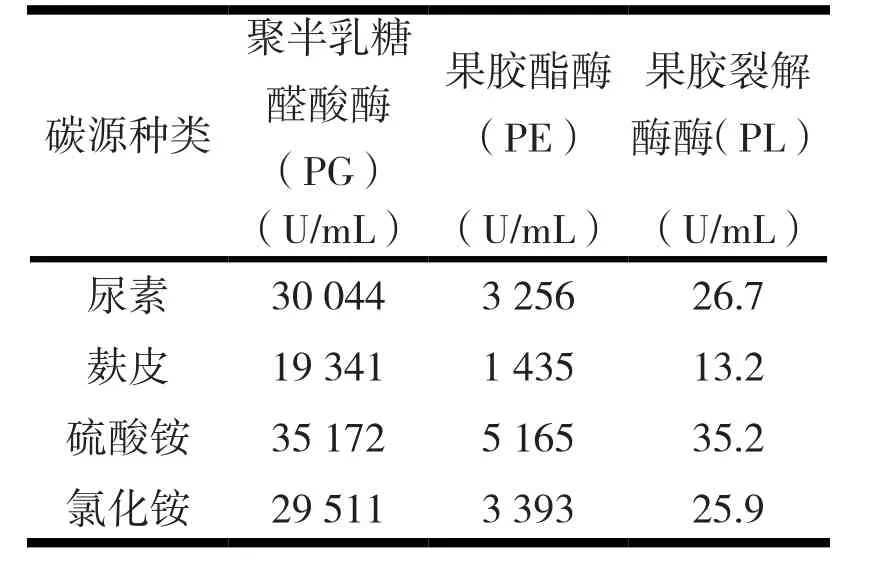
表5 不同氮源对诱变株Y-26产酶活力的影响
由表5可知,无机氮源硫酸铵产酶活力最高,平均达到35 172 U;尿素为氮源时酶活力次之;麦麸为氮源,菌株Y-26产酶力最低。因此,Y-26菌株以硫酸铵为氮源进行研究。
2.3.3 不同金属离子与诱变株Y-26产酶活力。金属离子能促进或抑制许多酶的产生,本文以Fe2+、Mg2+、Ca2+的不同浓度探讨了诱变株Y-26的产酶活力。结果见表6。
表6结果表明,Mg2+对产酶影响最大,对产酶有一定的促进作用; Ca2+对产酶有一定的抑制作用; Fe2+对产酶基本无影响。
2.3.4 初始pH与诱变株Y-26产酶活力。在上述优化的基础上进行了不同起始pH对诱变株产酶活力的影响,结果见表7。
由表7可知,诱变株Y-26对pH的适应范围较广,培养基起始pH在3~8均可发酵产酶,当pH为5,诱变株Y-26产酶活力最高。

表6 不同金属离子对诱变株Y-26产酶活力的影响
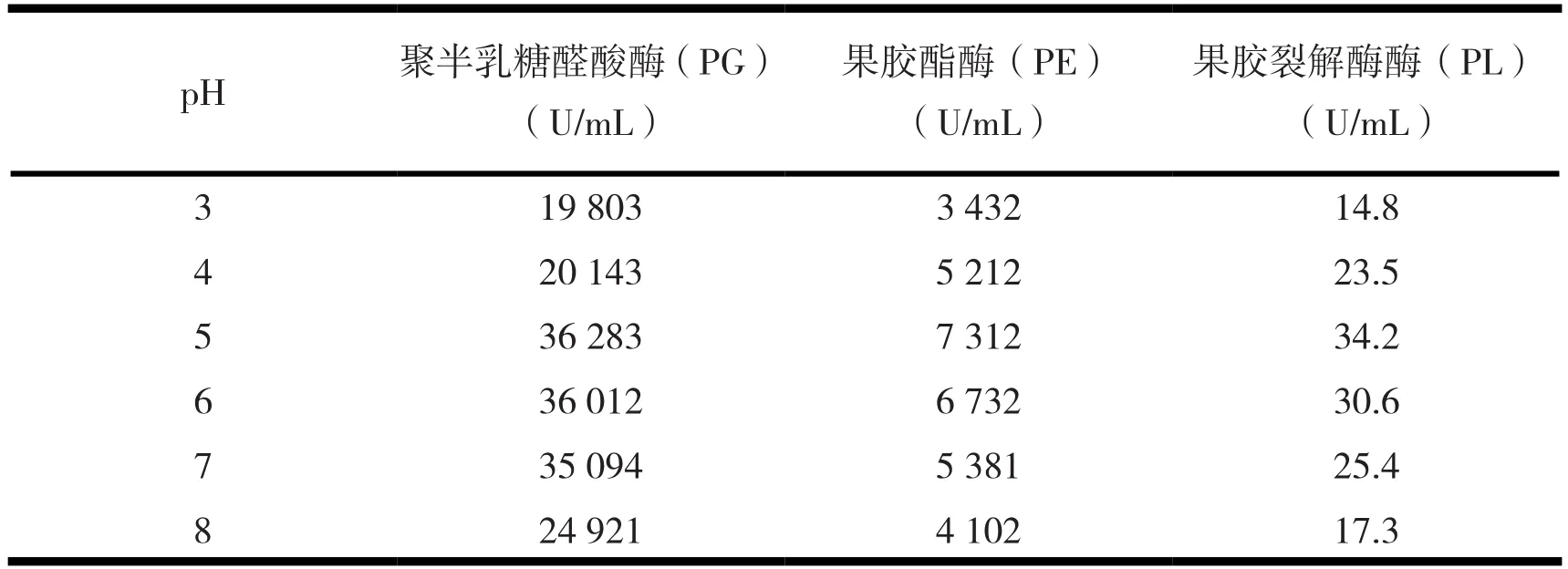
表7 初始pH对诱变株Y-26产酶活力的影响
2.3.5 培养时间与诱变株Y-26产酶活力。应用前面优化出的最佳培养条件,考察培养时间对诱变株Y-26产酶活力的影响。结果见表8。
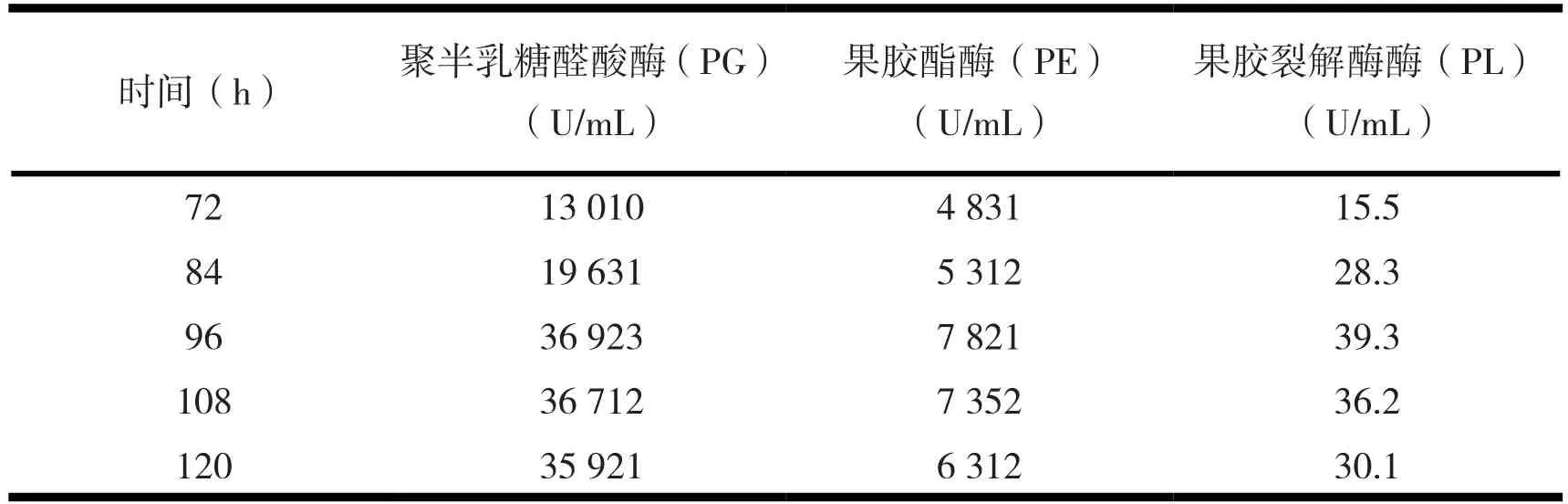
表8 培养时间对诱变株Y-26产酶活力的影响
由表8知,在时间为96~120 h范围内酶活力较高,酶活力没有显著差异。考虑到产酶活力的稳定性、节省时间及发酵所需的能源,最佳发酵产酶时间有待进一步通过正交试验优化。
2.3.6 不同温度与诱变株Y-26产酶活力。诱变株Y-26置于不同温度中,实验不同温度与诱变株Y-26产酶活力,结果见表9。
由表9知,在温度为30℃~40℃范围内酶活力较高,考虑到节省能源,选择30℃为最佳发酵产酶温度。

表9 培养温度对诱变株Y-26产酶活力的影响
2.4 诱变株Y-26酶制剂脱囊衣工艺初步研究
以酶制剂用量(酶浓度/%)、料液比(g/mL)、处理时间(min)、酸浓度(%)、温度(℃)为实验因素,以砂囊得率为酶解囊衣效果评价指标,采用正交实验法对发酵酶制剂脱囊衣工艺进行优化,实验因素设计与水平见表10。
由表10可知,根据R值大小为E>D>B>C>A,即影响酶解脐橙囊衣得率的主次顺序为温度>酸浓度>料液比>酶浓度>时间;由表中K值分析可知酶解脱囊衣的最优工艺组合为A4B1C3D4E4,即酶解时间为120 min,料液比为1:2,酶添加浓度为5%,酸浓度为0.5‰,酶解温度为50 ℃。因此,需要以该工艺条件为验证试验,最后的砂囊得率为48.4%,故经验证实验,A4B1C3D4E4为酶制剂脱囊衣的最佳工艺条件。

表10 正交实验设计因素与水平表
3 结语
用传统筛选和紫外诱变育种相结合技术手段,构建遗传稳定、产酶活力高且对脐橙囊衣降解具有高度专一性的微生物——卷枝毛霉。初步确定脐橙脱囊衣专用产酶卷枝毛霉的发酵生产工艺条件,为工业化大规模生产该酶制剂提供理论参考。用该工艺生产的产品,不添加任何防腐剂,延长产品保质期,原有的酸碱脱囊衣技术将被彻底更新换代。通过改善脐橙橙汁胞加工工艺,促进资源节约和环境保护,降低生产成本,产品更具竞争力,产品质量更安全可靠,有利于出口。
猜你喜欢
昆明医科大学学报(2021年5期)2021-07-22
中华养生保健(2020年3期)2020-11-16
World Journal of Clinical Cases(2019年6期)2019-04-17
猪业科学(2018年5期)2018-07-17
山东工业技术(2016年15期)2016-12-01
广东饲料(2016年2期)2016-12-01
中国酿造(2016年12期)2016-03-01
中国卫生标准管理(2015年3期)2016-01-14
应用海洋学学报(2015年3期)2015-11-22
化工进展(2015年6期)2015-11-13
