
基于宏基因策略的新颖木聚糖酶基因cbxynA克隆及其重组酶酶学性质
2019-10-12 06:01崔晓亭王小艺赵峙尧柳佳芸裴鹏刚熊苏玥
中国食品学报 2019年9期
熊 科 崔晓亭 王小艺 赵峙尧 柳佳芸 裴鹏刚 邓 蕾 熊苏玥
(1北京工商大学 北京市食品添加剂工程技术研究中心 北京 100048
2北京工商大学 北京食品营养与人类健康高精尖创新中心 北京 100048
3北京工商大学 北京市质量与安全重点实验室 北京 100048
4北京工商大学 北京市风味化学重点实验室 北京 100048)
我国是农业大国,伴随着农产品生产加工,每年产生大量农业废弃物。这些农业废弃物资源再生利用率极低,大部分被浪费[1-2]。大力开发利用这些资源可以缓解环境、资源和粮食危机,因此农业废弃物再利用成为近十几年各国研究的热点[3]。木聚糖是结构比较复杂的半纤维素,为世界第二大可再生资源[4]。由于木聚糖结构复杂,它的彻底水解需要一系列酶的协同作用,其中最关键的酶是木聚糖酶[5]。在众多应用中,以木聚糖经酶法制取有较高附加值的功能低聚糖,是富含半纤维素材料的农业废弃物再生利用的有效增值途径,具有巨大的潜在经济效益和环保意义[6-8]。而木聚糖酶属水解酶类,在碳循环过程中发挥了不可替代的功能,具有重要的潜在应用价值,目前已成功应用于食品、饲料、酿造、造纸、医药和能源等领域[9-11]。
目前,多数木聚糖酶来源于各类天然微生物菌株,通过天然产酶方式获得的木聚糖酶多为复合酶,表达量低,且不易纯化[12]。由于自然界中99%以上的微生物是不能被纯培养利用,因此该方法得到的产木聚糖酶菌株十分有限,仅局限于不到1%的微生物基因资源,对自然界中所占比例较大的非培养微生物资源无法利用[13]。近年来,越来越多的学者基于宏基因组的方法筛选木聚糖酶。如汪国增等[14]通过构建瘤胃宏基因组文库,从中筛选到酶学性质较为新颖的木聚糖酶,其最适作用温度30℃,可用于果汁的澄清,生产饲料等工业。张鹏等[15]从堆肥构建的宏基因组文库中筛选到性质较为优良的纤维素酶和木聚糖酶,可用于造纸等工业。Bao等[16]从牦牛瘤胃等宏基因文库中筛选到能提高木聚糖酶活力的纤维素酶,可用于食品加工,动物饲料的加工。Hu Y等[17]从土壤宏基因文库中筛选到一种低温耐碱酶,与常规木聚糖酶不同,可用于制备生物燃料、燃料乙醇。GonX等[18]从牛瘤胃等中筛选到无纤维素酶活力的木聚糖酶,可用于纸浆漂白等工业化应用。Mo XC等[19]从土壤宏基因组文库中筛选到一种多功能酶,能够制备低聚木糖。从来源上比较,木聚糖酶广泛来源于各种环境,如海洋、陆地等。而土壤微生物是木聚糖酶的巨大来源库,通常每1 g土壤中有几亿到几百亿个微生物,虽然土壤宏基因文库构建面临的干扰因素多、难点大,如腐殖酸的去除等,但构建土壤未培养微生物的宏基因组文库能更广泛地获得木聚糖酶优良基因资源。本研究以土壤环境作为宏基因来源对酶进行发掘,相比其它环境的宏基因来源可大大提高寻找和发现新木聚糖酶基因的概率,理论意义和潜在的应用价值较大。利用HiTAIL-PCR将获得非培养的木聚糖酶基因全长进行异源表达,为更好地开发利用土壤未培养微生物资源,发掘新颖的木聚糖酶资源奠定基础。
1 材料与方法
1.1 样品采集
土壤样品来自云南、贵州、河南、河北等15个地区。样品采用铁铲采集,用刮刀除去5 cm厚的表层土。将采集的土壤样品保存于无菌塑料袋中,于-4℃冰箱保存待用。
1.2 主要试剂
DNA琼脂糖凝胶回收试剂盒(美国OMEGA);琼脂糖(西班牙 Biowest);十六烷基三甲基溴化铵(CTAB)、十二烷基磺酸钠(SDS)购自上海生工生物技术有限公司;RNA酶,蛋白酶K来自(日本 TAKARA);Sau 3A I、BamH I、CIAP、T4 DNA 连 接 酶 、Q5 DNA 聚 合 酶 、λDNA/Hind III、DL2000、1 kb PlusDNA Ladder,pUC19、E.coli DH5、pET28a 载体、E.coli BL21(DE3)感受态细胞(日本TAKARA);氨苄霉素(Amp)蛋白胨(Tryptone)、酵母提取物(Yeast Extract)(英国 Oxford公司);Gel Exraction Kit(美国 OMEGA);刚果红,卡那霉素(美国 AMRESCO);榉木木聚糖(美国 Sigma)。
1.3 主要设备与仪器
DYC P-31BN水平电泳槽、DYY-10C电泳仪,北京六一仪器厂;Image Quant 300凝胶成像仪、Multitemp III恒温循环水浴器,美国 GE公司;Sigma 1-14小型台式高速离心机,美国 Sigma公司;WD-9405B型水平摇床,北京六一;PCR仪,Biometra,德国;ImageQuant 300 凝胶成像仪,Multitemp III恒温循环水浴器,美国GE;琼脂糖凝胶电泳仪,北京六一;DHZ-DA全温振荡器,江苏太仓培英;WD-9405B型水平摇床,北京六一;Nanodrop核酸定量仪,Thermo公司。
1.4 土壤宏基因组文库构建
每次称取80 g土壤,用提取缓冲液(Tris-HCl:100 mmol/L pH 8.0;EDTA:100 mmol/L,pH 8.0;NaCl:1.5 mol/L;CTAB:1%)溶解稀释土样,225 r/min。混匀2~3 h。具体提取方法参照Zhou等[20]。将回收后的DNA片段采用Sau3A I进行部分酶切,并与经BamH I酶切后与pUC19载体进行连接,电转化法导入到DH5α中。
1.5 土壤宏基因组文库的筛选
土壤宏基因组文库的筛选方法参照Kwon等[21],将文库中的克隆子转接到含有0.5%的木聚糖底物的平板上培养24 h,将平板放置在50℃烘箱30 min后,刚果红染色15 min,1 mol/L的Na-Cl脱色15 min,在底物平板上能产生透明圈的克隆即为阳性克隆。
1.6 木聚糖酶基因cbxynA序列分析
对上述筛选得到的阳性克隆子提取质粒并测序,用在线软件预测外源插入片段所有可能的开放阅读框,用DNAMAN预测目的基因可能的分子质量和等电点。
1.7 重组酶CBXYNA的表达和纯化
利用Primer5软件设计引物,扩增cbxynA的ORF编码序列。在序列 N端正(5’-CATGCCATGGCACACGTACACGTCCCG-3’)引入Xho I位点,在C端(5’-CCGCTGAGGCAGGCTGCGAAAAGCCC-3’)引入 Nco I位点。
将目的基因cbxynA经Xho I和Nco I双酶切后连接到pET-28a(+)载体并转化到BL21(DE3)感受态细胞中进行异源表达。对表达过程中何时加入IPTG的诱导时机、加入IPTG诱导后的温度、诱导的时间及IPTG浓度等参数进行优化。收集菌体后用裂解液裂解,超声波破碎细胞后离心收集上清粗酶液。用Ni柱纯化粗酶液,先用2.5%wash&elution buffer对柱料进行冲洗,通过A280检测器对除杂蛋白情况进行监控,直至检测器显示没有杂峰的出现。用wash&elution buffer对柱料进行梯度洗脱,梯度分别为5%,10%,20%,40%,60%,80%,100%。并将对应的出峰时间的蛋白进行收集。用SDS-PAGE电泳检测纯化后的酶蛋白。
1.8 重组酶CBXYNA活力及酶学性质测定
以木聚糖为底物,采用DNS法测定酶活力。酶最适pH值和pH稳定性测定缓冲液体系为pH值2.2~11。每种缓冲体系5个点,共35个点。重组CBXYNA酶活力的测定在50℃下进行,将常规方法中的缓冲液替换为上述不同pH体系的缓冲液,测定所得酶活力记为100%。pH稳定性是将酶液稀释,使酶液处于不同的缓冲体系中,50℃下保温30 min后立即冰浴终止反应,之后按照DNS测定方法测定残余酶活力,以未经保温处理的重组木聚糖酶CBXYNA活力为100%,分别计算不同pH下的残余相对酶活力。
测定最适温度时,用最适pH缓冲液将酶稀释一定比例,然后置于40~80℃的不同温度下,反应10 min,测定酶活力,以最大值为100%。温度稳定性的测定,将样品与酶最适反应缓冲溶液以一定比例混匀,置于不同温度(50,55,60,65,70 ℃)中保温不同时间(0,30 min,1 h,1.5 h,2 h,3 h,4 h,5 h)。定时取样,测定样品残留酶活力,以未处理木聚糖酶活力作对照。
考察的金属离子 Na+、K+、Li+、Ca2+、Mg2+、Zn2+、Mn2+、Fe3+、Al3+、Cu2+对 CBXYNA 酶活力的影响。酶对底物特异性的研究,添加1%(w/v)的燕麦木聚糖、桦木木聚糖、水溶性玉米芯木聚糖、水不溶性玉米芯木聚糖在0.05 mol/L,pH 5.2的醋酸缓冲液中,反应时间为10 min,反应温度为55℃条件下测定木聚糖酶活力。
2 结果与分析
2.1 土壤宏基因组文库的构建及其筛选
从土壤中提取总的DNA,经纯化浓缩后的23 kb以上的DNA的量约为6 μg。因土壤环境中得到的总DNA片段较长,不利于外源片段插入载体中。故采用Sau3A I酶对回收的土壤DNA片段进行部分酶切,再用1%的琼脂糖凝胶回收2~10 kb以内的DNA片段约2 μg左右。土壤粗提DNA电泳图如图1所示。
将酶切时间设置为1 h,每100 μL加入0.75 U的Sau3A I酶,使得土壤DNA的条带分布在2~8 kb,符合构建质粒土壤宏基因组文库。酶切之后的土壤DNA与pUC19载体经T4连接酶连接后,导入到DH5α中。将转化产物涂布平板后获得了大量宏基因文库克隆子。从文库中随机提取10个克隆子并提取质粒,用EcoR I和Hind III双酶切之后,结果表明:质粒间插入的外源酶切带型差异显著,文库克隆片段随机性较高。外源插入大小为1.5~4.5 kb,平均为 3 kb,粗略计算,宏基因组的库容量为30 Mb。
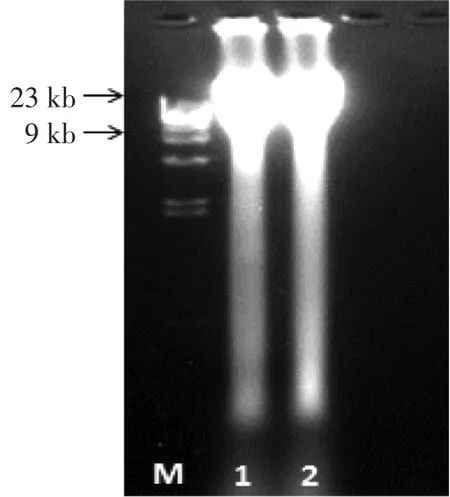
图1 不同方法提取土壤DNA电泳图Fig.1 Extraction and purification of DNA from soil by different methods
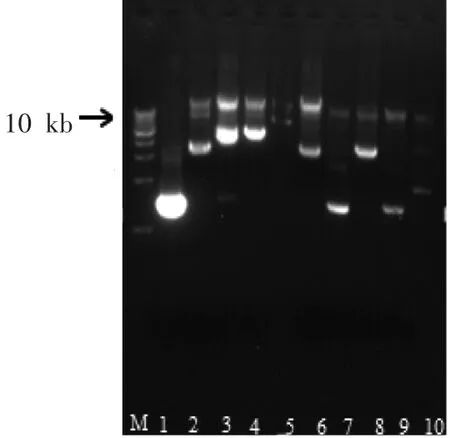
图2 土壤宏基因组文库重组质粒双酶切分析Fig.2 The restriction analysis of recombinant plasmids from the soil metagenomic library
随后,基于功能策略筛选,采用刚果红法对文库进行初筛,Cb5545克隆子在刚果红平板上的透明圈如图3所示。

图3 Cb5545克隆子初筛(a)及复筛(b)选平板中的透明圈Fig.3 Transparent rings of Cb5545 clone in primary screening(a)and screening(b)plate
平板筛选结果表明:克隆子Cb5545具有水解木聚糖底物的活性,使以木聚糖为底物的平板经刚果红染色后产生了阳性透明圈。随后对筛选的克隆子进行摇瓶发酵,取粗酶液采用DNS法测定发现其初始酶活力为12.09 U/mL。
2.2 基因cbxynA序列分析
将筛选得到的阳性克隆子Cb5545测序,得到一段4 238 bp的DNA序列,并预测了该外源插入片段的所有可能开放阅读框,其中开放阅读框ORF10是一个822 bp长的编码木聚糖酶的完整的开放阅读框,其编码产物的氨基酸序列与来自Halobacteriaceae archaeon(古细菌属)编码的木聚糖酶的相似度最高也仅为45%,表明该序列是一条未知的新序列(GenBank登记号:KY062986)。该基因编码的产物由273个氨基酸组成,预测的分子质量为29.1 ku,等电点为5.2。
2.3 重组酶CBXYNA原核表达优化
将目的基因cbxynA克隆到pET-28a(+)表达系统里进行异源表达。对重组质粒何时加入IPTG诱导的时机、加入IPTG后的诱导时间、诱导温度及IPTG浓度进行优化。最终的优化结果如图4所示。结果表明:培养5 h后,添加IPTG终浓度为0.7 mmol/L,在23℃下诱导25 h,获得的最大酶活力为52 U/mL,较初始值提高了4.3倍。
2.4 重组酶CBXYNA的纯化
CBXYNA经硫酸铵盐析和Ni-NTA纯化结果如表1所示。硫酸铵盐析和Ni-NTA纯化后的产物经SDS-PAGE验证(图5泳道2),将得到的电泳纯级CBXYNA木聚糖酶进行酶学特性研究。
2.5 重组酶CBXYNA酶学性质研究


图4 IPTG诱导时机的菌体生长曲线(a)及IPTG加入后的诱导时间(b)、诱导温度(c)及诱导剂浓度(d)对CBXYNA酶活力影响Fig.4 Growth curve of muton for IPTG inducing(a)and Effect of IPTG inducing time(b),temperature(c),concentration(d)on CBXYNA enzyme activity
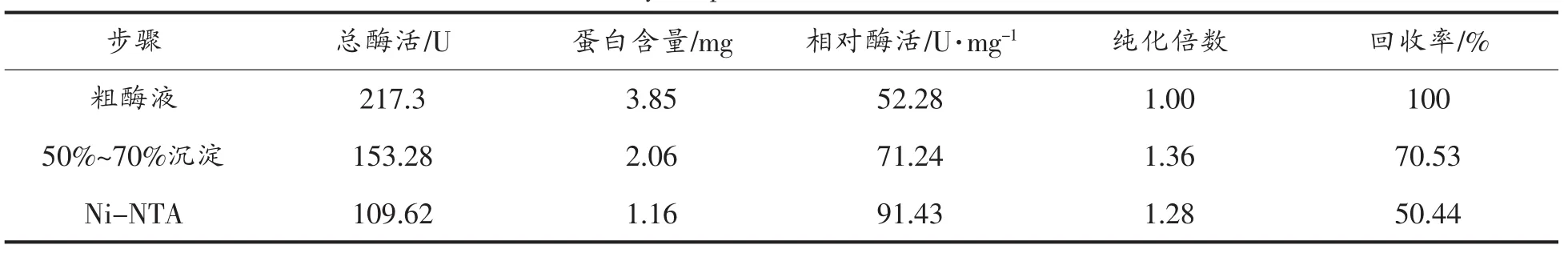
表1 重组木聚糖酶CBXYNA的纯化总结Table 1 Summary of purification recombinant CBXYNA
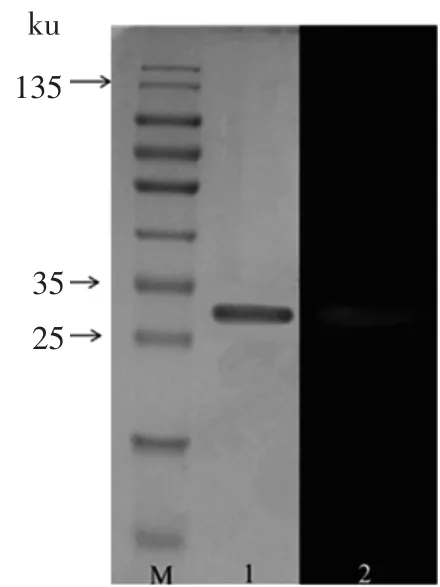
图5 CBXYNA纯化过程的电泳图和酶谱图Fig.5 The SDS-PAGE electrophoresis and zymogram of recombinant CBXYNA
当以木聚糖为底物时,CBXYNA酶促反应的最适pH为5.2(图6a),其pH稳定性在pH 4.2到10.2之间,保温30 min后残余酶活仍能达到70%以上(图6b)。CBXYNA的酶促反应最适温度为55℃(图6c),在50~60℃条件下保温30 min,残余酶活力仍为80%以上(图6d)。
研究 Ca2+、Fe3+、Cu2+、Mn2+、Mg2+、Zn2+、K+和Na+、等不同金属离子对重组木聚糖酶CBXYNA酶活力的影响,由图7可知,Na+、 K+、Ca2+、Li+在1~7 mmol/L的范围之内对酶有促进作用,而Mg2+则在1~3 mmol/L内对酶有促进作用,在5~7 mmol/L对酶有抑制作用。Mn2+、Zn2+、Fe3+、Al3+、Cu2+对酶有抑制作用,且抑制作用随着金属离子浓度的升高而增强。其中,Cu2+对酶的抑制作用最明显,几乎使其完全丧失酶活。金属离子在微生物机体内主要是作为酶活性中心的组成部分,维持生物大分子细胞结构的稳定性。但是有些金属离子对机体有不利影响,研究认为半胱氨酸可以参与糖残基-酶中间体的形成,而有些重金属离子阻碍了这一过程,不利于酶促反应的进行,也可能是影响了酶与底物的结合及解离状态。然而金属离子影响酶活力的具体机制还有待于进一步的研究。
进一步探究CBXYNA酶的底物特异性,以桦木木聚糖的酶活力为对照100%,当以水溶性玉米芯木聚糖作为底物时,CBXYNA酶活力达到162%;以燕麦木聚糖为底物时,酶活力为139%。以水不溶性玉米芯为底物时,酶活力为74%;以麸皮为底物时,其酶活力为56%(表2)。
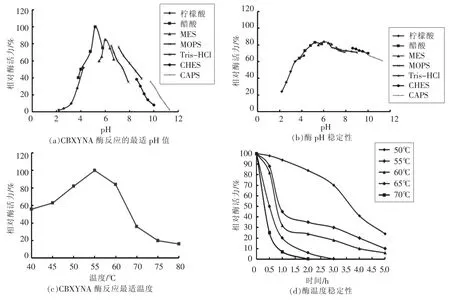
图6 重组酶CBXYNA的酶学性质Fig.6 Enzymatic properties of the recombinant enzyme CBXYNA

图7 金属离子对木聚糖酶活性的影响Fig.7 Effect of metal Ions on xylanase activity
3 结论
研究从土壤宏基因文库中筛选并鉴定出一个木聚糖酶基因cbxynA,经分析表明该基因是一段未知的新序列。随后对该基因进行克隆表达,对其原核表达条件进行了优化,最终获得其最大酶活力为52 U/mL,较初始值提高了4.3倍。重组木聚糖酶 CBXYNA,其最适 pH为 5.2,pH在4.2到10.2之间时其酶活力在70%以上,有较好的pH稳定性。CBXYNA的最适温度为55℃,在50~60℃保温30 min酶活力仍在80%以上,具较好的热稳定性。Na+、K+、Ca2+、Li+在1~7 mmol/L 的范围之内对酶有促进作用,Mn2+、Zn2+、Fe3+、Al3+、Cu2+对酶有抑制作用,且抑制作用随着金属离子浓度的升高而增强。其中,Cu2+对酶的抑制作用最明显,几乎使其完全丧失酶活。当该酶以水溶性玉米芯木聚糖、燕麦木聚糖、水不溶性玉米芯木聚糖、酶活力分别为162%,139%,74%,说明该CBXYNA更容易结合水解水溶性木聚糖底物。重组木聚糖酶CBXYNA有较好的pH稳定性,可用于造纸、饲料、食品等工业应用中。同时该酶以燕麦木聚糖、水溶性玉米芯木聚糖为底物时,酶活力相对较高,有较强的亲和能力,可被用于水解可溶性木聚糖底物制备低聚木糖。
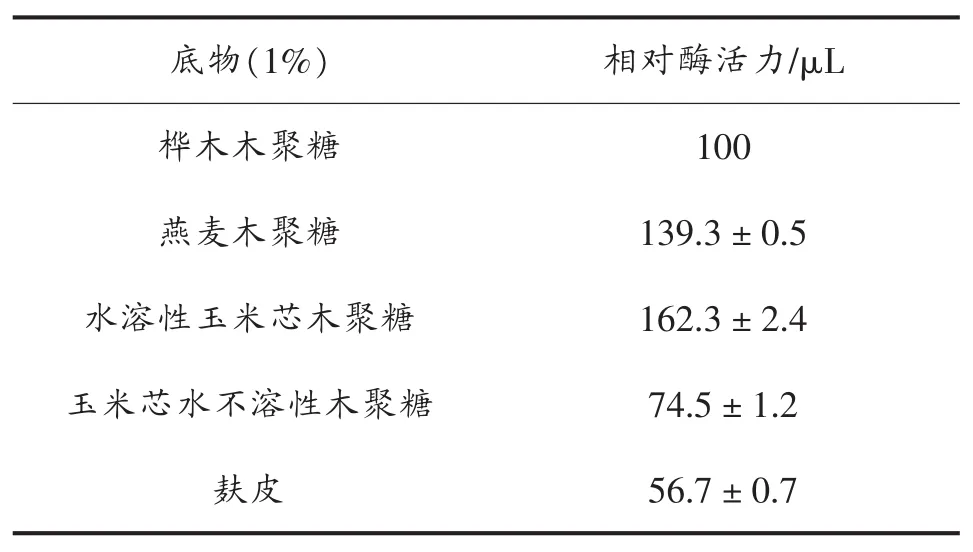
表2 重组CBXYNA对不同水解底物的特异性Table 2 Specificity of CBXYNA towards different hydrolysis substrates
猜你喜欢
湘潮(上半月)(2022年7期)2022-12-06
动物营养学报(2022年9期)2022-10-12
粮食加工(2021年5期)2021-11-01
猪业科学(2021年3期)2021-05-21
中国饲料(2021年2期)2021-02-23
幽默大师(2020年10期)2020-11-10
教育周报·教育论坛(2020年3期)2020-10-21
科学(2020年2期)2020-08-24
中华诗词(2019年1期)2019-11-14
湖南饲料(2019年5期)2019-10-15
