
退化喀斯特森林群落常见植物叶片光合作用变异特征
2019-11-04 06:38罗绪强张桂玲王世杰邓家彬刘兴阮英慧杨成
生态环境学报 2019年9期
罗绪强 ,张桂玲, ,王世杰,邓家彬,刘兴 ,阮英慧,杨成
1. 贵州师范学院地理与资源科学学院,贵州 贵阳 550018;2. 中国科学院普定喀斯特生态系统观测研究站,贵州 普定 562100;3. 贵州师范学院化学与材料学院,贵州 贵阳 550018;4. 贵州民族大学生态环境工程学院,贵州 贵阳 550025
光合作用是自然界实现能量与物质转化的最重要的化学反应(匡经舸等,2017),是植物干物质积累的基础和最重要的生理过程,对植物生长发育有着重要的意义(韩忠明等,2014;李明月等,2013)。植物光合作用与陆地生态系统碳水循环密不可分,是一个内外因子共同作用的复杂过程(翟占伟等,2017;Smith et al.,2000),通常是净光合速率(Pn)、蒸腾速率(Tr)、气孔导度(Gs)、胞间CO2浓度(Ci)、光合有效辐射(PAR)、冠层结构、空气相对湿度、大气相对温度、大气CO2浓度等诸多生态因子相互作用的综合结果(铁军等,2018;Deng et al.,2000)。其中,Pn、Tr、Gs和 Ci是植物光合作用在生理上形成的对环境适应机制的体现(杨玉珍等,2018;薛雪等,2015;罗绪强等,2013),是表征陆地生态系统植物光合作用最直接的4个气体交换参数,一定程度上,它们能够反映植物在生长区的生存和竞争能力(李征珍等,2017;云建英等,2006)。目前,有关植物光合作用及其影响因素方面的研究已有大量报道,但主要侧重单个因子的影响机制或多种因子复合影响的模拟研究(Nowicka et al.,2018;高冠龙等,2018;韩吉梅等,2017;鲍芳等,2017;Erel et al.,2015;Xu,2001),对自然生态系统特别是喀斯特森林生态系统植物群落演替过程中植物光合作用对环境变化响应方面的研究还鲜见报道。
喀斯特森林是在喀斯特地貌上生育的森林,因其生境的特殊性,结构的复杂性和系统的脆弱性,在一系列特征上与常态地貌上生育的森林有明显不同(Zeng et al.,2007;侯满福等,2006)。喀斯特森林植被群落结构相对简单,自调控能力较弱(罗绪强等,2014;刘映良等,2005),它只在环境参数严格限定之内才能持续稳定存在,若环境参数或种群值受到重大扰乱,系统将面临崩溃而发生逆转,使生境的严酷性剧增甚至植物难以生存,最终形成石漠化景观(邓晓琪等,2012;郭柯等,2011;Wang et al.,2004;Yuan,1997)。退化喀斯特森林是结构和功能退化并失去固有平衡的森林生态系统,是喀斯特森林由顶极群落到草本群落逆向演替的过程(罗绪强等,2018;喻理飞等,2000)。本研究选取陈旗小流域为研究区,以流域内退化喀斯特森林群落中的常见植物为研究对象,分别对其叶片净光合速率(Pn)、蒸腾速率(Tr)、气孔导度(Gs)、胞间CO2浓度(Ci)等气体交换参数进行测定分析,探讨了不同演替阶段群落植物叶片气体交换参数对环境变化的响应,以期为喀斯特森林生态系统的稳定性维护和退化生态系统的修复提供数据支撑和理论依据。
1 材料与方法
1.1 研究区概况
陈 旗 小 流 域 ( 26°15′36″- 26°15′56″N ,105°43′30″-105°44′42″E)位于贵州省安顺市普定县境内,地跨陈旗堡和赵家田两村,属长江流域的乌江水系,是中国科学院普定喀斯特生态系统观测研究站监测研究区。该流域面积1.29 km2,下伏基岩为三叠系中统关岭组灰岩和泥灰岩,土壤以石灰土为主,土层稀少、浅薄、分布不连续。区内地形起伏大,山坡陡峻,海拔1338-1491 m。属亚热带季风湿润气候区,多年平均降水量1336 mm,年蒸发量920 mm,年均气温14.2 ℃,7月最热,平均气温22.6 ℃,1月最冷,平均气温4.1 ℃。具有贵州典型的喀斯特峰丛山体、峰丛洼地地貌以及喀斯特水文地质特征和山地土地利用方式。流域内有不同演替阶段的植物群落,从稀灌草丛、藤刺灌丛、灌木林、乔灌过渡林到乔木林,构成了具有代表性的完整演替序列,主要物种有火棘(Pyracantha fortuneana)、粉枝莓(Rubus biflorus)、小叶鼠李(Rhamnus parvifolia)、红叶木姜子(Litsea rubescens)、小果蔷薇(Rosa cymosa)、竹叶花椒(Zanthoxylum armatum)、大披针薹草(Carex lanceolata)、中国蕨(Sinopteris grevilleoides)、柏木(Cupressus funebris)、响叶杨(Populus adenopoda)等。
1.2 研究对象
在流域内选取稀灌草丛、藤刺灌丛、灌木林、乔灌过渡林和乔木林5个演替阶段植物群落内均常见的火棘、竹叶花椒、小叶鼠李和小果蔷薇共4种植物为研究对象。其中,火棘是蔷薇科(Rosaceae)火棘属(Pyracantha)常绿灌木,竹叶花椒是芸香科(Rutaceae)花椒属(Zanthoxylum)落叶小乔木,小叶鼠李是鼠李科(Rhamnaceae)鼠李属(Rhamnus)灌木,小果蔷薇是蔷薇科(Rosaceae)蔷薇属(Rosa)攀援灌木。
1.3 实验设计
在稀灌草丛、藤刺灌丛和灌木林群落中分别选取200 m2,在乔灌过渡林和乔木林群落中分别选取300 m2的样方作为实验样地,在不同样地内均选取树龄相当(根据株高和胸径进行估算)的火棘、竹叶花椒、小叶鼠李和小果蔷薇各3株进行植物叶片光合作用参数的测定,每个样地测定12株,5个样地共测定 60株。用于光合作用参数测定的所有样本都选自同一朝向(向阳方向)且高度相当的当年生完全展开的成熟、健康叶片,测定前轻拭植物叶片上的尘土或其它杂质,测定时保持叶片自然生长角度不变,同一叶片重复测量5次,每株植物测量3片叶后取平均值代表该植株测定结果。
1.4 测定方法
2017年7月(植物生长季)28-30日,在天气晴朗无风的上午8:00-11:00,采用美国LI-COR公司生产的 LI-6400便携式光合测定系统分析仪(LI-COR,Lincoln,USA)测定植物叶片光合作用气体交换参数。以CO2注入系统为气源,控制叶室内 CO2浓度为接近于大气 CO2的浓度 400 μmol·mol-1;光合有效辐射(PAR)设定为 1000 μmol·m-2·s-1,由 LI-6400-02B 型红蓝光源提供;叶室温度设定为 25 ℃。测定植物叶片的净光合速率(Pn,μmol·m-2·s-1,以 CO2计)、蒸腾速率(Tr,mmol·m-2·s-1,以 H2O 计)、气孔导度(Gs,mol·m-2·s-1,以 H2O 计)和胞间 CO2浓度(Ci,µmol·mol-1,以 CO2计)等光合参数。水分利用效率(WUE)=Pn/Tr(µmol CO2/mmol H2O);叶片羧化速率(CE)=Pn/Ci(mol·m-2·s-1);气孔限制值(Ls)=1-Ci/C0,C0为 400 µmol·mol-1。
1.5 数据分析
用Microsoft Excel 2010进行数据整理,用IBM SPSS Statistics 19进行统计分析。统计分析包括描述性统计分析(Descriptive Statistics)、单因素方差分析(Compare Means)和多因素方差分析(General Linear Model)及相关分析(Correlate)等。
2 结果与分析
2.1 流域内植物叶片光合参数的总体特征
从表1可以看出,研究区常见植物叶片的Pn、Tr、Gs和 Ci分别在 1.13-22.40、0.13-7.62、0.02-0.64和 75.00-369.00之间变化;平均值分别为9.95、2.71、0.22和 269.88。其中,Ci的最大值与最小值之比和变异系数均最小,但方差和标准差均最大,说明流域内植物叶片的Ci相对其他3个光合参数来说最稳定,但离散程度最高。
植物叶片的 WUE、CE和 Ls分别在 0.91-30.15、3.06-86.84和 0.08-0.81之间变化;平均值分别为 5.04、37.57和 0.33。其中,Ls的最大值与最小值之比、变异系数、方差和标准差均最小,说明流域内植物叶片的气孔限制值相对水分利用效率和羧化速率来说,是最稳定且离散程度也是最低的。
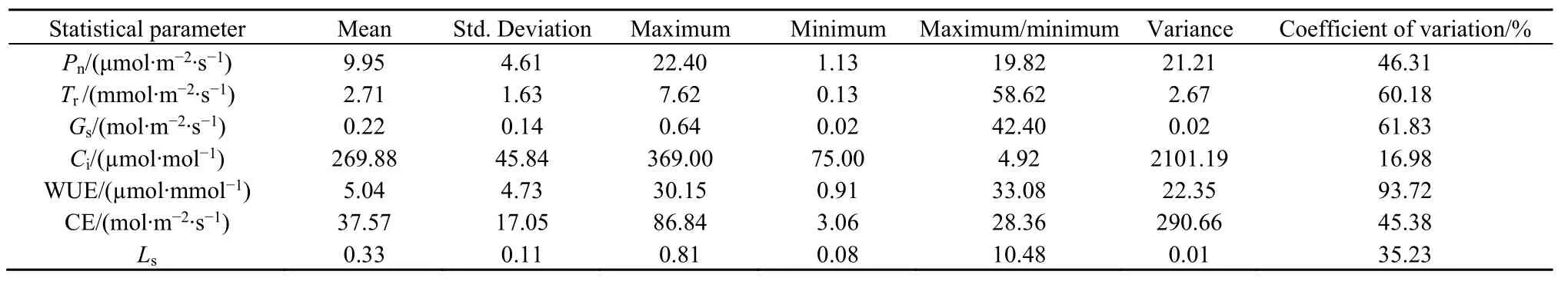
表1 研究区常见植物叶片的光合参数、水分利用效率、羧化速率和气孔限制值Table 1 Photosynthetic parameters, water use efficiency, carboxylic rate and stomatal limitation value in leaves of common plants in the study site
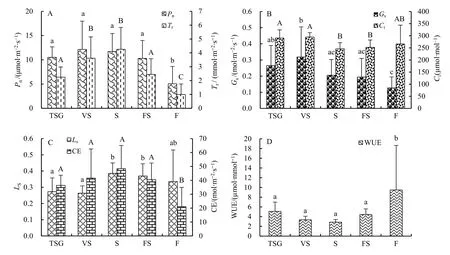
图1 不同群落植物叶片的Pn-Tr(A)、Gs-Ci(B)、Ls-CE(C)、WUE(D)值Fig. 1 Pn-Tr (A), Gs-Ci (B), Ls-CE (C), and WUE (D) of leaves of different community plants
2.2 植物叶片光合参数的群落变异特征
从图1可以看出,研究区各演替阶段植物群落常见植物叶片 Pn的平均值表现为藤刺灌丛(12.14)>灌木林(11.72)>稀灌草丛(10.51)>乔灌过渡林(10.29)>乔木林(5.07),方差分析结果表明群落间差异显著(P=0.000),多重比较后按同类子集可将其分为乔灌过渡林+稀灌草丛+灌木林+藤刺灌丛(P=0.304)群落和乔木林群落共 2类。Tr的平均值表现为灌木林(4.26)>藤刺灌丛(3.61)>乔灌过渡林(2.45)>稀灌草丛(2.25)>乔木林(1.00),群落间差异显著(P=0.000),按同类子集可将其分为乔木林群落、稀灌草丛+乔灌过渡林(P=0.694)群落、藤刺灌丛+灌木林(P=0.199)群落共3类。Gs的平均值表现为藤刺灌丛(0.32)>稀灌草丛(0.27)>灌木林(0.21)>乔灌过渡林(0.19)>乔木林(0.13),差异显著(P=0.006),按同类子集分为乔木林+乔灌过渡林+灌木林(P=0.153)群落、乔灌过渡林+灌木林+稀灌草丛(P=0.195)群落、稀灌草丛+藤刺灌丛(P=0.300)群落共3类。Ci的平均值表现为藤刺灌丛(294.67)>稀灌草丛(290.75)>乔木林(266.00)>乔灌过渡林(252.25)>灌木林(245.75),差异显著(P=0.019),按同类子集分为灌木林+乔灌过渡林+乔木林(P=0.280)群落、乔木林+稀灌草丛+藤刺灌丛(P=0.126)群落共2类。统计分析结果表明,除乔木林群落外,各群落常见植物叶片Pn、Tr、Gs、Ci的最大值与最小值之比和变异系数均较小,说明植物叶片各光合参数在乔木林群落外的同一演替阶段植物群落中均较稳定。Pn、Tr、Gs在乔木林群落中的最大值与最小值之比均大于10且变异系数均大于50%,说明植物叶片Pn、Tr、Gs在乔木林群落内部均不稳定,变异程度都较高。
各演替阶段植物群落常见植物叶片 Ls的平均值表现为灌木林(0.39)>乔灌过渡林(0.37)>乔木林(0.34)>稀灌草丛(0.27)>藤刺灌丛(0.26),方差分析结果表明群落间差异显著(P=0.019),多重比较后按同类子集可将其分为藤刺灌丛+稀灌草丛+乔木林(P=0.126)群落、乔木林+乔灌过渡林+灌木林(P=0.280)群落共2类。CE的平均值表现为灌木林(48.34)>藤刺灌丛(41.61)>乔灌过渡林(40.45)>稀灌草丛(36.37)>乔木林(21.06),群落间差异显著(P=0.001),按同类子集分为乔木林群落和稀灌草丛+乔灌过渡林+藤刺灌丛+灌木林(P=0.076)群落共 2类。WUE的平均值表现为乔木林(9.48)>稀灌草丛(5.09)>乔灌过渡林(4.44)>藤刺灌丛(3.35)>灌木林(2.86),差异显著(P=0.003),按同类子集分为灌木林+藤刺灌丛+乔灌过渡林+稀灌草丛(P=0.247)群落和乔木林群落共2类。统计分析结果表明,除乔木林群落外,各群落常见植物叶片Ls、CE、WUE的最大值与最小值之比和变异系数均较小,说明植物叶片的气孔限制值、羧化速率和水分利用效率在乔木林群落外的同一演替阶段植物群落中均较稳定。在乔木林群落中,Ls、CE、WUE与 Pn、Tr、Gs一样,最大值与最小值之比均大于10且变异系数均大于50%,说明乔木林群落内的植物叶片Ls、CE、WUE同样不稳定,变异程度也都较高。
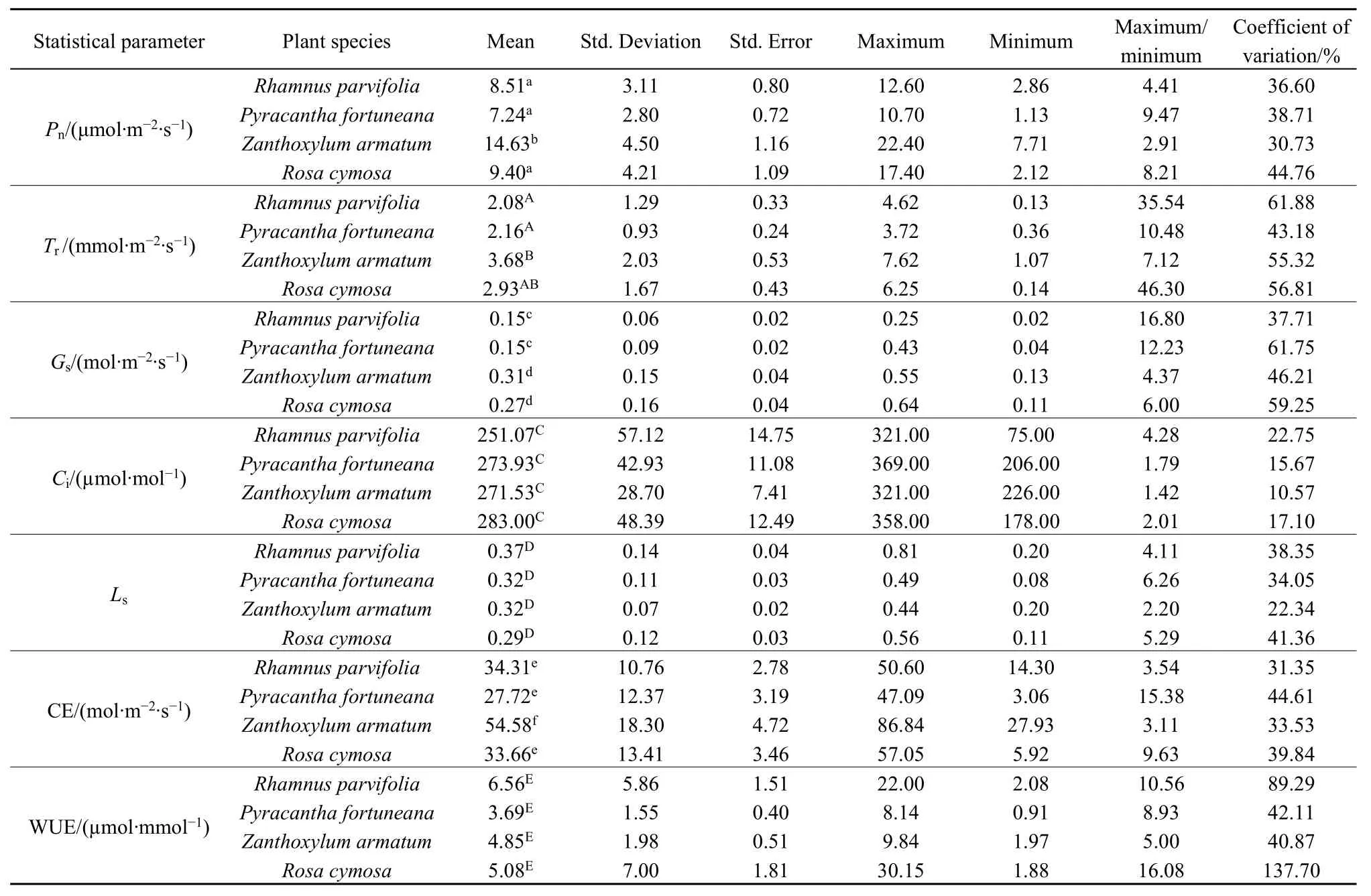
表2 流域内不同种植物叶片的光合参数、水分利用效率、羧化速率和气孔限制值Table 2 Leaf photosynthetic parameters, water use efficiency, carboxylic rate and stomatal limitation value of different plant species in the study site
2.3 植物叶片光合参数的种间差异特征
2.3.1 流域内的种间差异
从表 2可以看出,研究区常见植物叶片的 Pn以竹叶花椒(14.63)最大,火棘(7.24)最小,方差分析表明种间差异显著(P=0.000),多重比较后按同类子集分为火棘+小叶鼠李+小果蔷薇(P=0.138)和竹叶花椒2类。Tr以竹叶花椒(3.68)最大,小叶鼠李(2.08)最小,种间差异显著(P=0.020),按同类子集分为小叶鼠李+火棘+小果蔷薇(P=0.158)和小果蔷薇+竹叶花椒(P=0.189)2类。Gs以竹叶花椒(0.31)最大,火棘(0.15)最小,种间差异显著(P=0.001),按同类子集分为火棘+小叶鼠李(P=0.978)和小果蔷薇+竹叶花椒(P=0.278)2类。Ci以小果蔷薇(283.00)最大,小叶鼠李(251.07)最小,种间差异不显著(P=0.277)。
植物叶片Ls以小叶鼠李(0.37)最大,小果蔷薇(0.29)最小,种间无显著差异(P=0.277)。CE以竹叶花椒(54.58)最大,火棘(27.72)最小,种间差异显著(P=0.000),按同类子集分为火棘+小果蔷薇+小叶鼠李(P=0.230)和竹叶花椒2类。研究区植物叶片 WUE以小叶鼠李(6.56)最大,火棘(3.69)最小,种间差异不显著(P=0.429)。
从表2还可以看出,研究区常见植物除小叶鼠李、小果蔷薇叶片的Tr和WUE、火棘叶片的Gs的最大值与最小值之比均大于 10且变异系数均大于50%以外,其余植物叶片的Pn、Tr、Gs、Ci和Ls、CE、WUE的最大值与最小值之比和变异系数均较小,说明在流域层面上,这些植物的叶片光合参数及相关指标在物种内均较稳定。
2.3.2 群落内的种间差异
从图2A可以看出,稀灌草丛中的植物叶片Pn以小果蔷薇(12.10)最大,火棘(8.36)最小,种间差异不显著(P=0.113)。藤刺灌丛中以竹叶花椒(19.53)最大,小叶鼠李(6.21)最小,种间差异显著(P=0.000),按同类子集分为小叶鼠李+火棘(P=0.294)、小果蔷薇、竹叶花椒共3类。灌木林中以竹叶花椒(17.17)最大,火棘(9.34)最小,种间差异显著(P=0.003),按同类子集分为火棘+小果蔷薇+小叶鼠李(P=0.427)、竹叶花椒共 2类。乔灌过渡林以竹叶花椒(15.03)最大,小果蔷薇(7.11)最小,种间差异显著(P=0.006),按同类子集分为小果蔷薇+火棘+小叶鼠李(P=0.050)、竹叶花椒共2类。乔木林以竹叶花椒(9.80)最大,火棘(2.32)最小,种间差异显著(P=0.018),按同类子集分为火棘+小果蔷薇+小叶鼠李(P=0.285)、竹叶花椒共2类。
从图2B可以看出,稀灌草丛中植物叶片Tr以小果蔷薇(2.74)最大,小叶鼠李(1.73)最小,种间差异不显著(P=0.207)。藤刺灌丛中以小果蔷薇(5.22)最大,小叶鼠李(1.90)最小,种间差异显著(P=0.004),按同类子集分为小叶鼠李+火棘(P=0.201)、竹叶花椒+小果蔷薇(P=0.317)共2类。灌木林中以竹叶花椒(6.43)最大,火棘(2.62)最小,种间差异显著(P=0.002),按同类子集分为火棘+小果蔷薇(P=0.104)、小果蔷薇+小叶鼠李(P=0.519)、竹叶花椒共3类。乔灌过渡林以竹叶花椒(4.06)最大,小果蔷薇(1.79)最小,种间差异显著(P=0.011),按同类子集分为小果蔷薇+火棘+小叶鼠李(P=0.753)、竹叶花椒共 2类。乔木林以竹叶花椒(1.51)最大,小叶鼠李(0.59)最小,种间差异不显著(P=0.509)。
从图2C可以看出,稀灌草丛中植物叶片Gs以小果蔷薇(0.38)最大,小叶鼠李(0.14)最小,种间差异不显著(P=0.119)。藤刺灌丛中以竹叶花椒(0.48)最大,小叶鼠李(0.14)最小,种间差异显著(P=0.002),按同类子集分为小叶鼠李+火棘(P=0.645)、小果蔷薇+竹叶花椒(P=0.990)共2类。灌木林中以竹叶花椒(0.34)最大,火棘(0.11)最小,种间差异显著(P=0.007),按同类子集分为火棘+小果蔷薇+小叶鼠李(P=0.113)、竹叶花椒共2类。乔灌过渡林以竹叶花椒(0.35)最大,小果蔷薇(0.12)最小,种间差异显著(P=0.031),按同类子集分为小果蔷薇+火棘+小叶鼠李(P=0.467)、竹叶花椒共 2类。乔木林以小果蔷薇(0.17)最大,火棘(0.07)最小,种间差异不显著(P=0.399)。
从图2D可以看出,稀灌草丛中植物叶片Ci最大的是小果蔷薇(312.00),最小的是小叶鼠李(265.00),种间差异不显著(P=0.393)。藤刺灌丛中以小果蔷薇(313.33)最大,竹叶花椒(279.67)最小,种间差异显著(P=0.045),按同类子集分为竹叶花椒+火棘+小叶鼠李(P=0.083)、小叶鼠李+小果蔷薇(P=0.326)共2类。灌木林中以小果蔷薇(264.67)最大,火棘(215.00)最小,种间无显著差异(P=0.058)。乔灌过渡林以竹叶花椒(271.67)最大,小果蔷薇(210.67)最小,种间差异显著(P=0.017),按同类子集分为小果蔷薇、火棘+小叶鼠李+竹叶花椒(P=0.598)共2类。乔木林以小果蔷薇(314.33)最大,小叶鼠李(167.00)最小,种间差异显著(P=0.041),按同类子集分为小叶鼠李、竹叶花椒+火棘+小果蔷薇(P=0.433)共2类。
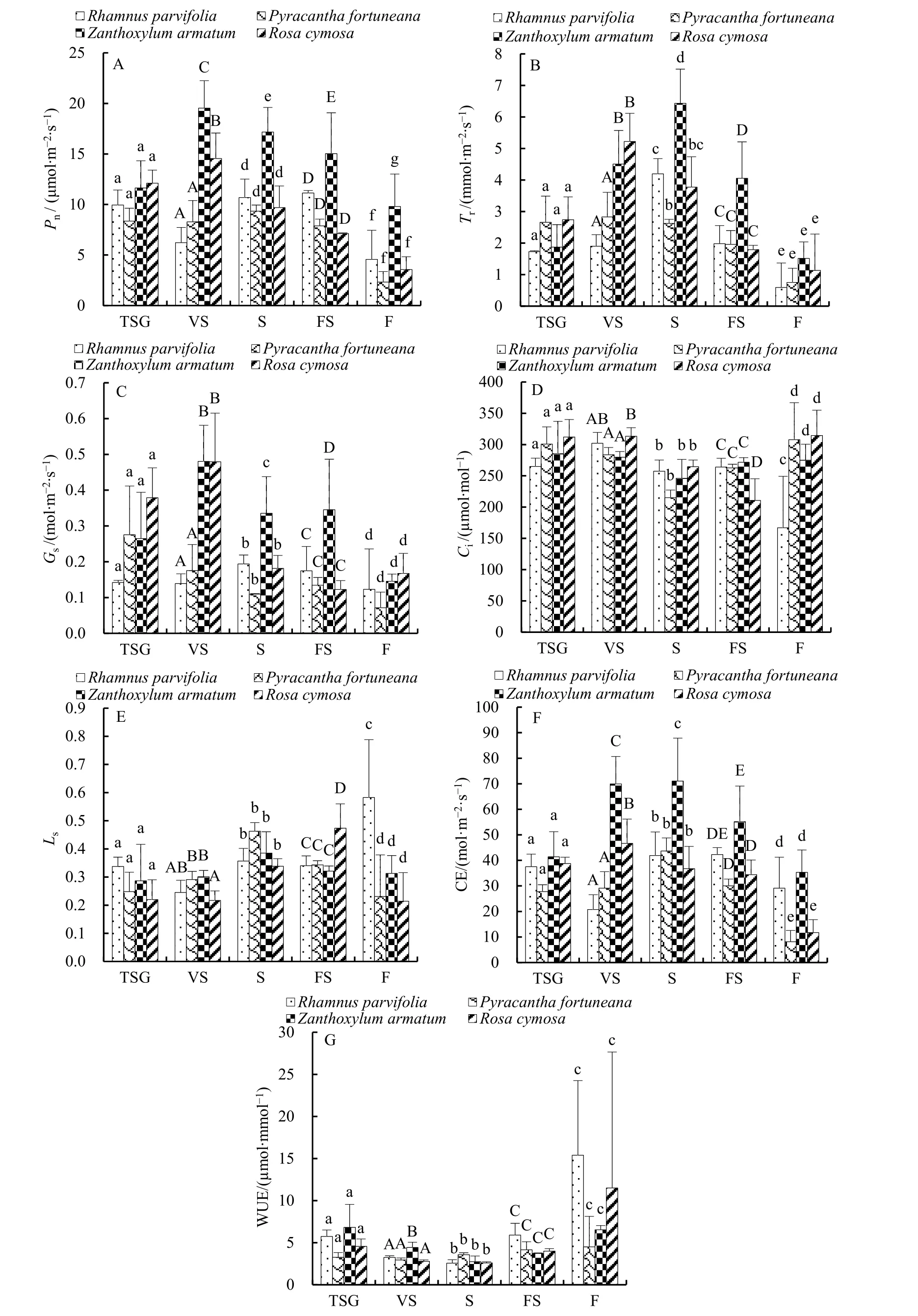
图2 不同群落各植物叶片的Pn(A)、Tr(B)、Gs(C)、Ci(D)、Ls(E)、CE(F)、WUE(G)值Fig. 2 Pn (A), Tr (B), Gs (C), Ci (D), Ls (E), CE (F), and WUE (G) of different plant species in different communities
从图2E可以看出,稀灌草丛中植物叶片Ls以小叶鼠李(0.34)最大,小果蔷薇(0.22)最小,种间无显著差异(P=0.393)。藤刺灌丛中以竹叶花椒(0.30)最大,小果蔷薇(0.22)最小,种间差异显著(P=0.045),按同类子集分为小果蔷薇+小叶鼠李(P=0.326)、小叶鼠李+火棘+竹叶花椒(P=0.083)共3类。灌木林中以火棘(0.46)最大,小果蔷薇(0.34)最小,种间差异不显著(P=0.058)。乔灌过渡林以小果蔷薇(0.47)最大,竹叶花椒(0.32)最小,种间差异显著(P=0.017),按同类子集分为竹叶花椒+小叶鼠李+火棘(P=0.598)、小果蔷薇共2类。乔木林以小叶鼠李(0.58)最大,小果蔷薇(0.21)最小,种间差异显著(P=0.041),按同类子集分为小果蔷薇+火棘+竹叶花椒(P=0.433)、小叶鼠李共2类。
从图2F可以看出,稀灌草丛中植物叶片CE以竹叶花椒(41.43)最大,火棘(27.76)最小,种间差异不显著(P=0.084)。藤刺灌丛中以竹叶花椒(69.95)最大,小叶鼠李(20.76)最小,种间差异显著(P=0.000),按同类子集分为小叶鼠李+火棘(P=0.258)、小果蔷薇、竹叶花椒共3类。灌木林中以竹叶花椒(71.05)最大,小果蔷薇(36.75)最小,种间差异显著(P=0.019),按同类子集分为小果蔷薇+小叶鼠李+火棘(P=0.476)、竹叶花椒共 2类。乔灌过渡林以竹叶花椒(55.16)最大,火棘(30.00)最小,种间差异显著(P=0.019),按同类子集分为竹火棘+小果蔷薇+小叶鼠李(P=0.101)、小叶鼠李+竹叶花椒(P=0.076)共2类。乔木林以竹叶花椒(35.29)最大,火棘(8.10)最小,种间差异显著(P=0.009),按同类子集分为火棘+小果蔷薇(P=0.602)、小叶鼠李+竹叶花椒(P=0.384)共2类。
从图2G可以看出,稀灌草丛中植物叶片WUE最高的是竹叶花椒(6.82),最低的是火棘(3.26),种间差异不显著(P=0.090)。藤刺灌丛中以竹叶花椒(4.42)最大,小果蔷薇(2.79)最小,种间差异显著(P=0.002),按同类子集分为小果蔷薇+火棘+小叶鼠李(P=0.169)、竹叶花椒共 2类。灌木林中以火棘(3.56)最大,小叶鼠李(2.56)最小,种间差异不显著(P=0.056)。乔灌过渡林以小叶鼠李(5.89)最大,竹叶花椒(3.72)最小,种间差异不显著(P=0.055)。乔木林以小叶鼠李(15.39)最大,火棘(4.50)最小,种间无显著差异(P=0.518)。
2.4 植物叶片光合参数间的相关性分析
从表3可以看出,研究区植物叶片各光合参数之间具有极显著正相关关系(P<0.01)的有Pn-Tr、Pn-Gs、Pn-CE、Tr-Gs、Tr-CE、Gs-Ci、Gs-CE 和 Ls-WUE,具有极显著负相关关系(P<0.01)的有 Tr-WUE、Ci-WUE和Gs-Ls,另外,WUE与Pn之间也具有显著负相关关系(P<0.05)。研究区植物叶片各光合参数之间的这种相关性,表明它们之间存在着一定的平行和依赖关系,这与前人的研究结果一致(匡经舸等,2017;曾群英等,2015;智颖飙等,2015;罗绪强等,2013)。

表3 植物叶片的光合参数、水分利用效率、羧化速率和气孔限制值之间的皮尔逊相关系数Table 3 Pearson correlation coefficients between photosynthetic parameters, water use efficiency, carboxylic rate and stomatal limitation value in leaves of common plants in the study site
3 讨论
植物叶片的光合生理特征既受自身遗传因素的影响,也与外部环境条件有关(杨玉珍等,2018;高冠龙等,2018;李征珍等,2017;曾群英等,2015)。不同演替阶段植物群落的环境物理特征及辐射状况不同,诱导着各种生态因子发生改变,并形成一定的环境梯度(李生等,2019;罗绪强等,2018;李征珍等,2017;曾群英等,2015)。为了解研究区植物自身遗传因素和外部环境条件对叶片光合生理特征的交互影响情况,以植物种和群落类型为控制变量,植物叶片 Pn、Tr、Gs、Ci、Ls、CE、WUE等光合参数为观测变量分别建立固定效应的饱和模型进行多因素方差分析。结果表明,研究区常见植物叶片的 Pn、Tr、Gs、Ci、Ls和 CE受植物种和群落类型的交互影响均极显著(P<0.01)。从多因素方差模型对观测变量数据的总体拟合程度来看,该模型对 Pn、Tr、CE 数据的拟合程度均较好(R2>0.80,调整的 R2>0.75),但对 Gs、Ci、Ls、WUE 数据的拟合程度均不理想(R2<0.80,调整的 R2<0.75),说明研究区植物叶片的 Pn、Tr、CE主要受自身遗传因素和生境条件的影响,而Gs、Ci、Ls、WUE除受自身遗传因素和生境条件的影响外,还受到其他因素的影响。另外,群落类型、植物种对研究区植物叶片的Pn、Tr、Gs、CE均有显著影响(P<0.01)并产生强烈的交互作用(P<0.01),但植物叶片的Pn、Tr、Gs、CE在各演替阶段群落内部均较稳定,而在不同群落之间均有显著差异(P<0.05),说明研究区植物叶片 Pn、Tr、Gs、CE的变异受生境条件的影响更大,植物叶片 Pn、Tr、Gs、CE的变异特征是植物光合作用对外界环境变化响应的结果。
4 结论
研究区植物的光合生理特征受自身遗传特性及生境条件的影响,光合参数之间存在着一定的平行和依赖关系。植物叶片 Pn、Tr、Gs、Ci、Ls、CE、WUE在不同演替阶段植物群落间均有显著差异(P<0.05),Pn、Tr、Gs和CE在不同种间的差异也均显著(P<0.05)。Pn、Tr、Gs、Ci、Ls和 CE 受植物种和群落类型的交互影响均极显著(P<0.01)。研究区植物叶片 Pn、Tr、Gs、CE受生境条件的影响较大,且对环境变化敏感,其变异特征是植物光合作用对外界环境变化响应结果的综合体现。
猜你喜欢
河南师范大学学报(自然科学版)(2022年5期)2022-08-08
林业勘察设计(2022年1期)2022-07-25
林业勘查设计(2021年3期)2021-06-07
少儿美术(2020年9期)2020-11-05
山东林业科技(2019年3期)2019-07-30
陶山(2019年1期)2019-03-29
蔬菜(2018年5期)2018-05-17
东坡赤壁诗词(2018年1期)2018-03-31
创新作文(1-2年级)(2017年5期)2017-12-07
绿色科技(2015年10期)2016-01-15
