
寄生经历和寄主龄期对斑痣悬茧蜂寄生草地贪夜蛾幼虫效能的影响
2019-11-27 09:34周金成张柱亭尚丹
植物保护 2019年5期
关键词:生物防治
周金成 张柱亭 尚丹
摘要 草地贪夜蛾Spodoptera frugiperda (J.E.Smith)属鳞翅目灰翅夜蛾属,是新近入侵我国的重大危险性害虫。斑痣悬茧蜂已被证明对同属于灰翅夜蛾属的甜菜夜蛾和斜纹夜蛾的幼虫均具有较强的寄生能力。为探究短期驯化是否能有效改善斑痣悬茧蜂对草地贪夜蛾幼虫的寄生能力,本研究比较了有、无寄生经历的斑痣悬茧蜂雌蜂分别对2龄和3龄草地贪夜蛾幼虫在寄生行为和效能上的差异。结果表明,有寄生经历的雌蜂相对于无寄生经历的雌蜂能够较快刺扎寄主幼虫,并表现出更高的刺扎频率和寄生率,但雌蜂的各项行为指标在2龄和3龄幼虫间无显著差异。本研究将为改善本土天敌对入侵性草地贪夜蛾的防控能力提供借鉴。
关键词 入侵害虫; 寄生蜂; 孤雌产雌; 生物防治
中图分类号: S 476.3, S 433.4 文献标识码: A DOI: 10.16688/j.zwbh.2019388
Abstract Spodoptera frugiperda (J.E.Smith), belonging to the genus Spodoptera, Lepidoptera, is a new dangerous invasive pest in China. Meteorus pulchricornis has been demonstrated as an effective biological control agent against the larvae of Spodoptera litura and Spodoptera exigua, which are both belonging to the genus Spodoptera. To investigate whether the shortterm training is helpful to improve the parasitic efficacy of M. pulchricornis females against S. frugiperda larvae, we compared the parasitic behavior and efficacy of females with or without parasitic experience when they parasite 2nd instar and 3rd instar larvae of S. frugiperda. The results showed that the experienced females performed higher parasitism rate, more frequent sting behavior, and shorter time to parasite host larvae than inexperienced females, but the parasitic behaviors were not different between 2nd and 3rd instar larvae. The results will supply a reference for the improvement of native natural enemies for the control of invasive pests.
Key words invasive pest; parasitoid wasp; thelytoky; biological control
草地貪夜蛾Spodoptera frugiperda (J. E. Smith),又称秋黏虫,是一种原产于北美地区的多食性害虫[1],其已报道的寄主植物至少涵盖76个科,353种[2]。草地贪夜蛾自2019年1月首次报道入侵我国云南省以来,短时间内在我国迅速迁飞扩散至黄河流域以南的大部分省份。截至2019年8月1日,我国已有21个省份发现有草地贪夜蛾入侵,为害面积已超过60万 hm2[3-4]。综合现有情况,草地贪夜蛾入侵并在我国定殖及周年为害几成定局,仅依靠现有的应急防控手段短时间内完全扑杀入侵种群已无可能,对草地贪夜蛾的防控已演变为一场“持久战”[4-6]。
对草地贪夜蛾等入侵生物而言,入侵地本土生态控制因子的缺位通常是造成入侵物种快速扩张的重要原因。天敌逃逸假说(enemy release hypothesis)认为,入侵物种在入侵地的暴发基于以下两个根本原理: 1)原产地入侵物种具有专食性天敌而入侵地没有。2)入侵地本土天敌与入侵害虫缺乏协同进化历史,不能有效识别并取食入侵害虫,而只倾向于取食本土植食昆虫[7-8]。
因此,对草地贪夜蛾的长期防控应基于生态学原理,针对其特定生态位寻找有效的长效防控策略。考察和筛选专食性天敌来重新构建新的生态平衡是目前国内外控制入侵生物的主要策略之一。这一策略已取得了许多可供参考的成功案例。早在1888年,美国引进了澳洲瓢虫Rodolia cardinalis成功控制了入侵北美的一种原产于澳大利亚的吹绵蚧Icerya purchasi的暴发[8-9]。在我国,针对长期肆虐的入侵性害虫美国白蛾Hyphantria cunea,通过筛选和利用本土性白蛾周氏啮小蜂Chouioia cunea来抑制美国白蛾种群扩张已取得较好的效果[8]。然而,由于本土天敌缺乏同入侵害虫的协同进化历史,导致其在田间可能无法有效识别外来入侵物种。因此,需要对本土天敌进行必要的“驯化”,最终使其充分发挥对入侵性害虫的防控作用。例如通过使用目标害虫对专食性天敌进行长期饲养或者使天敌接触目标害虫的气味、寄主植物等,进而提高天敌对目标害虫的搜寻能力,改善其防控效果[10]。
斑痣悬茧蜂Meteorus pulchricornis是一种主要寄生鳞翅目夜蛾科害虫幼虫的寄生蜂。据报道,该蜂是甜菜夜蛾S.exigua和斜纹夜蛾S.litura幼虫的重要寄生性天敌,而这两种夜蛾科害虫与入侵我国的草地贪夜蛾同属于灰翅夜蛾属,具有较密切的亲缘关系[11-13]。基于此,我们认为斑痣悬茧蜂可能是防控草地贪夜蛾幼虫的一种重要候选天敌。由于4龄以上的草地贪夜蛾幼虫常具有钻蛀习性,而斑痣悬茧蜂尚未有报道具有可以钻入植株寄生鳞翅目幼虫的习性。因此,本研究分别选取了2龄和3龄草地贪夜蛾幼虫作为寄主,调查了无寄生经历和有寄生经历的两种雌蜂对2龄和3龄草地贪夜蛾幼虫的首次刺扎时间、刺扎频率、寄生率和结茧率。研究结果将为考察和筛选斑痣悬茧蜂防治草地贪夜蛾提供重要参考,同时也将为筛选和利用针对草地贪夜蛾的其他生防作用物提供借鉴。
1 材料与方法
1.1 供试昆虫
草地贪夜蛾幼虫采自贵州省黔东南州凯里市丹寨县附近的玉米田。野外采集的幼虫于室内以新鲜玉米叶饲养至成虫羽化后置于圆柱形养虫笼(直径20 cm,高35 cm)内产卵,笼壁覆盖纱布和牛皮纸作为草地贪夜蛾成虫产卵介质。用蘸有10%蜂蜜水的脱脂棉每12 h饲喂草地贪夜蛾成虫1次。待草地贪夜蛾卵块孵化后,幼虫统一以人工饲料饲养。人工饲料的配制主要以Bowling于1967年[14]提出的草地贪夜蛾人工饲料配方为基础,加入微量复合维生素、韦氏盐和甲醛加以改进。试验选取发育至2龄或3龄的草地贪夜蛾幼虫作为供试寄主。
本试验中的斑痣悬茧蜂室内种群由江苏科技大学生物技术学院提供。该蜂为孤雌产雌品系,即雌蜂无需交配即可产下100%的雌性子代。该蜂在室内以2龄末期的斜纹夜蛾幼虫为寄主已连续饲养20代以上。羽化后的雌蜂以10%蜂蜜水每日饲喂。试验统一选取2日龄雌蜂作为供试雌蜂。
1.2 试验设计
本试验预设2个试验因素,分别为寄主幼虫龄期和雌蜂是否具有寄生经历,共组成4组处理组合:即有寄生经历的雌蜂寄生2龄幼虫、有寄生经历的雌蜂寄生3龄幼虫、无寄生经历的雌蜂寄生2龄幼虫和无寄生经历的雌蜂寄生3龄幼虫。每组处理各设10次重复。
为获得有寄生经历的雌蜂,试验预先将20头草地贪夜蛾2龄末期幼虫置于底部平铺饲料的塑料养虫盒(长8 cm,宽4 cm,高4 cm)内,将1头雌蜂接入养虫盒内寄生,肉眼观察到雌蜂刺扎幼虫后,继续使雌蜂寄生24 h后移出,移出有寄生经历的雌蜂备用。为排除试验容器和接蜂操作造成的试验误差,将1头无寄生经历的雌蜂移入没有寄主幼虫的同一尺寸的养虫盒内24 h后,移出无寄生经历的雌蜂备用。试验前预先将每5头2龄幼虫或3龄幼虫分装于管壁平铺饲料的玻璃管(直径2 cm,长10 cm)内,将1头有寄生经历的雌蜂或无寄生经历的雌蜂接入玻璃管内寄生12 h后,移出寄生蜂。寄生蜂移入后2 h内观察统计雌蜂的首次刺扎时间、刺扎次数(产卵器成功刺入寄主虫体)。将幼虫单头分装于透气塑料盒内以人工饲料饲养至寄生蜂子代从虫体内钻出结茧,中途死亡的寄主幼虫解剖观察其体内是否含有寄生蜂幼虫。统计各重复内寄主幼虫的被寄生头数,记录寄生率和结茧率。
1.3 数据处理
采用广义线性模型下的双因素方差分析检验寄生经历和寄主龄期分别对雌蜂刺扎次数(泊松分布)、寄生率(二项分布)和子代蜂结茧率(二项分布)的影响。随接蜂时间的延长,雌蜂对寄主首次刺扎的累积风险采用考克斯比例风险模型进行分析。不同处理组合下各试验指标的多重比较采用Tukey 氏HSD法进行检验[15]。本试验所得数据均使用R统计软件进行分析[16]。
2 结果与分析
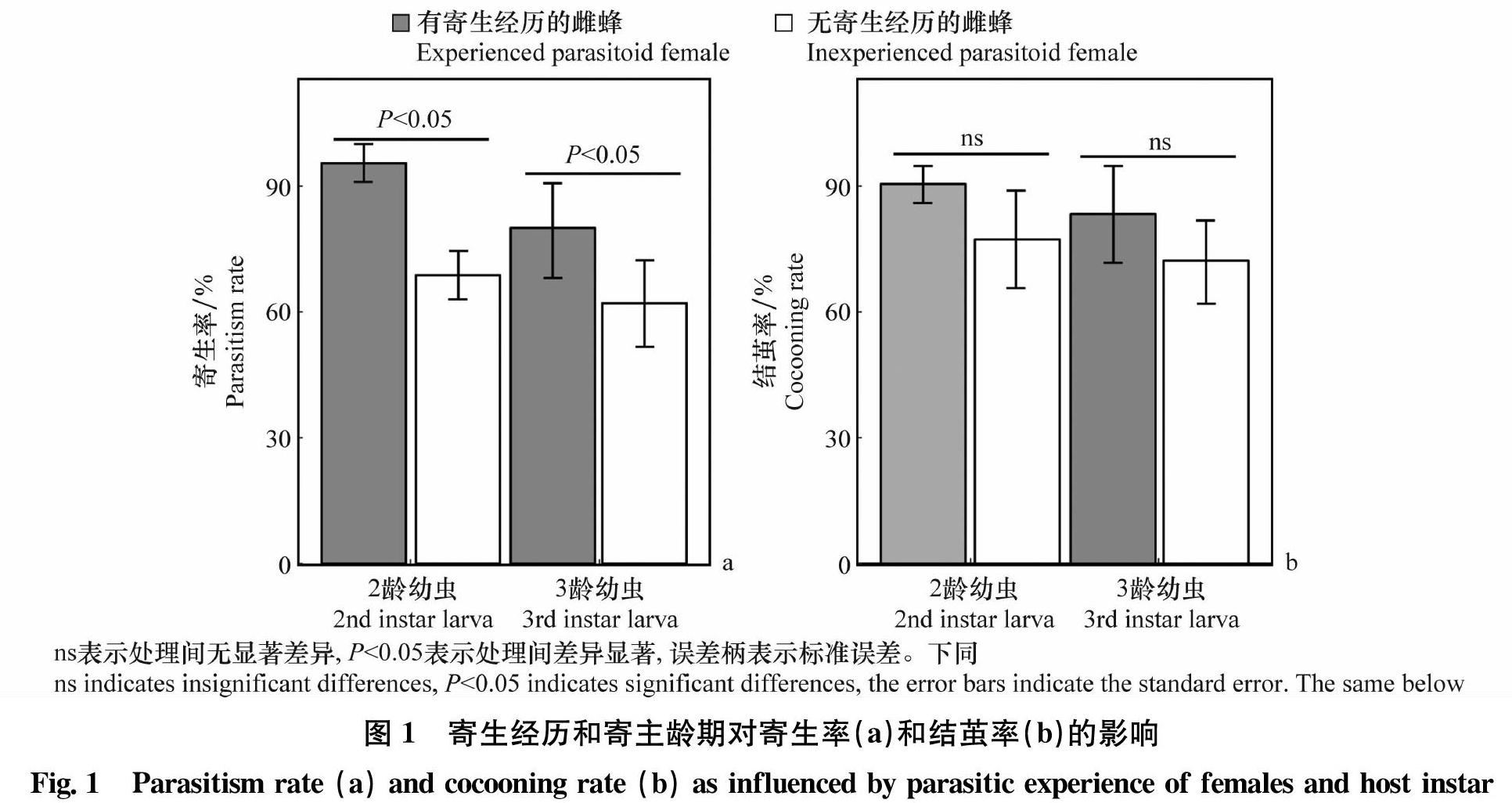
双因素方差分析表明,雌蜂寄生经历对寄主幼虫寄生率存在显著影响(F1,94=7.07,P<0.05)。有寄生經历的雌蜂的寄生率(90.16%±5.37%)显著高于无寄生经历的雌蜂(65.49%±6.11%;z=2.38,P<0.05)。寄主龄期(F1,94=1.28,P=0.26)和寄主龄期与寄生经历间的交互作用(F1,94=1.12,P=0.29)均对寄生率无显著影响(图1a)。子代蜂结茧率均不受雌蜂寄生经历(F1,69=1.75,P=0.11)、寄主龄期(F1,69=0.38,P=0.54)以及两因素间的交互作用(F1,69=0.078,P=0.78)的显著影响(图1b)。
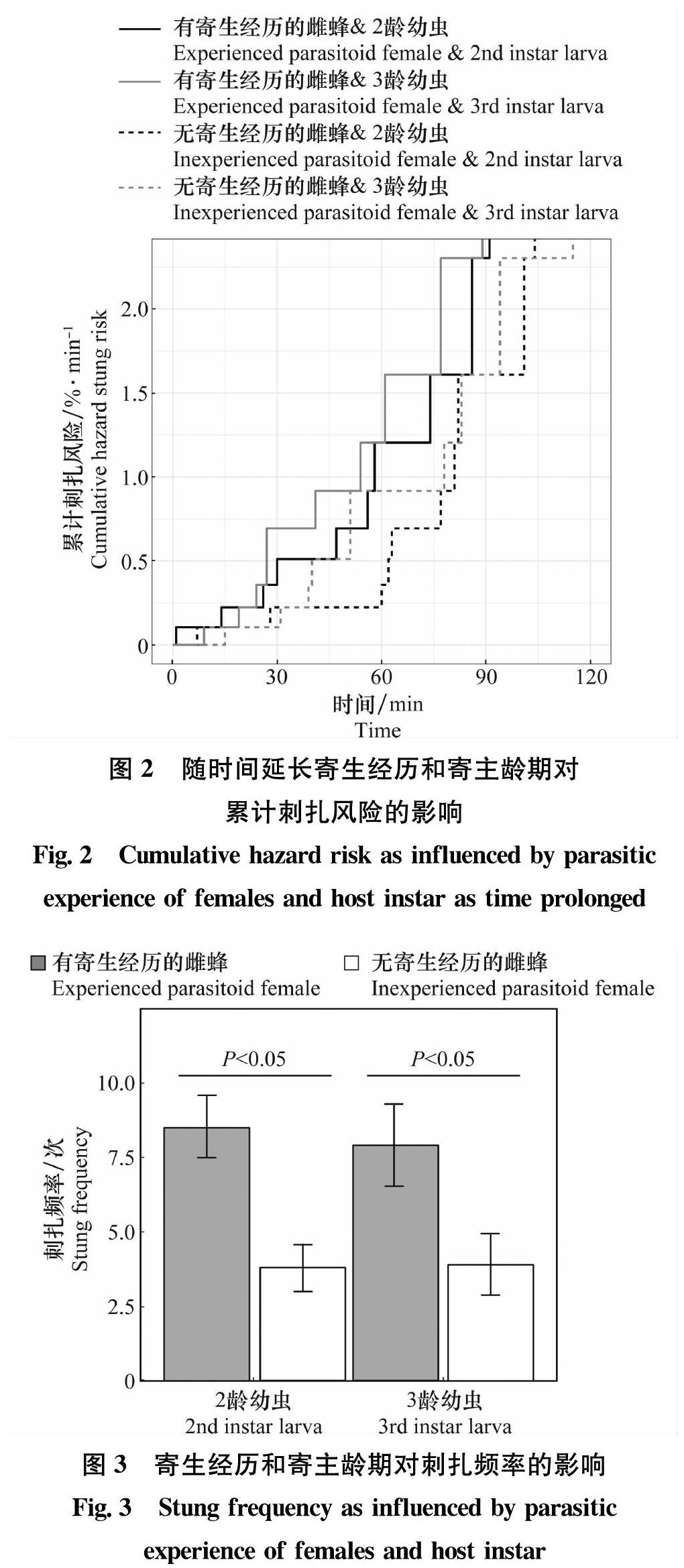
考克斯比例风险模型表明,雌蜂首次刺扎时间受雌蜂寄生经历的显著影响(χ2=4.02,df=1,P<0.05),但均不受寄主龄期(χ2=0.24,df=1,P=0.62)和寄生经历与寄主龄期间交互作用(χ2=0.074,df=1,P=0.79)的影响。随时间的延长,有寄生经历的雌蜂对寄主的瞬时刺扎风险(平均值±标准误差:1.41%/min±0.24%/min)显著高于无寄生经历的雌蜂(0.71%/min±0.12%/min;z=2.94,P<0.05)(图2)。有寄生经历的雌蜂首次刺扎寄主时的平均中位时间为44.01 min,而无寄生经历的雌蜂首次刺扎寄主的平均中位时间为62.52 min。
雌蜂刺扎频率受雌蜂寄生经历的显著影响(F1,36=13.16,P<0.05),但均不受寄主龄期(F1,36=0.043,P=0.84)和寄生经历与寄主龄期间交互作用(F1,36=0.053,P=0.82)的影响。有寄生经历的雌蜂对寄主的刺扎频率(8.19次±0.64次)显著高于没有寄生经历的雌蜂(3.85次±0.44次;z=5.47,P<0.05)(图3)。
3 讨论
研究结果表明,相对于无寄生经历的雌蜂,有寄生经历的斑痣悬茧蜂雌蜂完成对寄主的识别和刺扎产卵花费的时间更短,因而对寄主幼虫的寄生率更高。草地贪夜蛾为新近入侵我国的外来物种,与我国本土天敌类群没有在自然界中形成长期的协同进化关系,因而本土天敌可能难以识别这一外来物种。因此,提高本土天敌对外来目标害虫的适应性和识别能力决定了本土天敌的防控潜能。前人研究表明,寄生蜂可以利用联系性学习来提高其对寄主害虫的搜索效率和寄生率[10,17]。例如接触过锈色粒肩天牛Apriona swainsoni幼虫的管氏肿腿蜂Sclerodermus guani雌蜂在再次搜索寄主时,会表现出对这种天牛幼虫的趋向性[18]。有寄生经历的广赤眼蜂Trichogramma evanescens雌蜂搜索寄主所花费的时间比无寄生经历的雌蜂明显缩短[19]。类似的研究也发现,有寄生经历的斑痣悬茧蜂可以有效识别健康寄主,从而避免发生过寄生行为[17]。这些研究和我们的研究结果均表明,通过对斑痣悬茧蜂成蜂进行短时驯化来提高其对草地贪夜蛾幼虫的寄生效率具有可行性。在该蜂的生产和应用过程中,除了对其成蜂进行驯化外,还可以通过以斜纹夜蛾、甜菜夜蛾等草地贪夜蛾近缘物种或直接以草地贪夜蛾幼虫为寄主对其进行多代驯化,以提高其对目标害虫的识别能力和选择趋向。在今后的研究中,应当考虑开展对本土天敌昆虫的驯化,以提高其对草地贪夜蛾的控害效果。
本研究还发现,斑痣悬茧蜂雌蜂对2龄和3龄草地贪夜蛾幼虫的寄生行为和寄生率均未表现出差异。前人研究表明,斑痣悬茧蜂倾向寄生斜纹夜蛾和甜菜夜蛾的中间龄期幼虫,1龄幼虫由于营养有限,导致子代蜂无法顺利完成发育,而较高龄期的幼虫虽具有丰富的营养,但其免疫功能较强[11]。尽管草地贪夜蛾与斜纹夜蛾和甜菜夜蛾同属于灰翅夜蛾属,但与后两者不同的是,草地贪夜蛾的高龄幼虫常表现出钻蛀性,因而防治草地贪夜蛾应坚持“防早防小”的基本原则。我们认为斑痣悬茧蜂对较低龄期的草地贪夜蛾幼虫的寄生作用可以在防治窗口期发挥一定的作用。
此外,分布于我国的斑痣悬茧蜂均为孤雌产雌生殖的种群,而欧洲地区分布的斑痣悬茧蜂多进行两性生殖。我国仅在1984年报道过1例疑似的斑痣悬茧蜂雄蜂野外个体[20]。孤雌产雌的特性将为这种寄生蜂的工厂化繁育和田间应用带来诸多优点:首先,释放于田间的孤雌产雌品系的雌蜂无需交配即可产出可育后代,因而不必花费时间去搜寻雄蜂交配,从而直接发挥防控作用,具有极强的野外定殖能力。其次,在室内大规模繁育过程中,两性生殖的寄生蜂常常需要利用大量寄主去繁育无寄生功能的雄蜂,用于与雌蜂交配,而孤雌产雌生殖的寄生蜂会充分利用有限的寄主幼虫资源育出100%的具有寄生功能的雌蜂,大大降低了生产成本[21]。第三,两性生殖的许多寄生蜂类群的性别决定机制为染色体单双倍体条件下的CSD性别决定机制,当寄生蜂在室内近亲繁育过程中,由于相关等位基因位点趋于纯合,常导致后期雄蜂比例上升,而孤雌产雌生殖的寄生蜂则规避了这一问题[22]。综合斑痣悬茧蜂的诸多优点,我们认为该蜂对于目前肆虐我国多地的重大入侵害虫草地贪夜蛾具有一定的防治潜力。关于斑痣悬茧蜂对草地贪夜蛾的控害潜力和寄生后生长发育表现,尚需开展更为详细、全面的研究。
参考文献
[1] TODD E L, POOLE R W. Keys and illustrations for the armyworm moths of the noctuid genus Spodoptera Guenée from the Western Hemisphere [J]. Annals of the Entomological Society of America, 1980, 73(6): 722-738.
[2] 吴秋琳, 姜玉英, 吴孔明. 草地贪夜蛾缅甸虫源迁入中国的路径分析[J]. 植物保护, 2019, 45(2): 1-6.
[3] 姜玉英, 刘杰, 朱晓明. 草地贪夜蛾侵入我国的发生动态和未来趋势分析[J]. 中国植保导刊, 2019, 39(2): 33-35.
[4] 王磊, 陈科伟, 陆永跃. 我国草地贪夜蛾入侵扩张动态与发生趋势预测[J/OL]. 环境昆虫学报,2019,41(4):683-694.
[5] 唐璞, 王知知, 吴琼, 等. 草地贪夜蛾的天敌资源及其生物防治中的应用[J]. 应用昆虫学报, 2019, 56(3): 370-381.
[6] 李志刚,吕欣,押玉柯,等.粤港两地田间发现夜蛾黑卵蜂与螟黄赤眼蜂寄生草地贪夜蛾[J/OL].环境昆虫学报,2019,41(4):760-765.
[7] LIU Hong, STILING P. Testing the enemy release hypothesis: a review and metaanalysis [J]. Biological Invasions,2006,8(7):1535-1545.
[8] 萬方浩, 侯有明, 蒋明星. 入侵生物学[M]. 北京:科学出版社, 2015.
[9] 荆英, 黄建, 黄蓬英. 有益瓢虫的生防利用研究概述[J]. 山西农业大学学报(自然科学版), 2002(4): 299-303.
[10] 刘树生, 江丽辉, 李月红. 寄生蜂成虫在寄主搜索过程中的学习行为[J]. 昆虫学报, 2003, 46(2): 228-236.
[11] 胡浩,孟玲,李保平.斑痣悬茧蜂对不同龄期斜纹夜蛾幼虫的寄生功能反应[J].中国生物防治学报,2015,31(2):176-180.
[12] 郭林芳, 孟玲, 李保平. 斑痣悬茧蜂寄生对甜菜夜蛾幼虫行为的影响[J]. 南京农业大学学报, 2010, 33(5): 71-74.
[13] 叶昕海,杨义,梅洋,等.草地贪夜蛾基因组注释及分析[J/OL].环境昆虫学报, 2019,41(4):706-717.
[14] BOWLING C C. Rearing of two lepidopterous pests of rice on a common artificial diet [J]. Annals of the Entomological Society of America, 1967, 60(6): 1215-1216.
[15] FARAWAY J J. Extending the linear model with R:Generalized linear, mixed effects and nonparametric regression models[M].Second edition.Boca Raton: CRC Press, 2016.
[16] R Core Team. R: A Language and Environment for Statistical Computing [EB/OL]. R Foundation for Statistical Computing, Vienna,Austria.[2019-03-11].https:∥www.rproject.org/.
[17] 张博,冯素芳,黄露,等.斑痣悬茧蜂的寄主辨别能力及其影响因素[J].昆虫学报,2011,54(12):1391-1398.
[18] 王晓红,杨忠岐,唐艳龙,等.两种肿腿蜂对锈色粒肩天牛虫粪及其寄主植物气味的学习行为[J].环境昆虫学报,2011,33(3):357-363.
[19] 李保平,刘小宁.学习对广赤眼蜂寻找寄主和接受寄主行为的影响[J].昆虫天敌,2002(4):154-158.
[20] 李琼芳.斑痣悬茧蜂产雌孤雌生殖的初步观察[J].昆虫知识,1984(2):82-83.
[21] LIU Quanquan, ZHANG Tongshu, LI Chunxue, et al. Decisionmaking in a bisexual line and a thelytokous Wolbachiainfected line of Trichogramma dendrolimi Matsumura (Hymenoptera: Trichogrammatidae) regarding behavior toward their hosts [J].Pest Management Science,2018,74(7):1720-1727.
[22] HEIMPEL G E, DE BOER J G. Sex determination in the Hymenoptera [J].Annual Review of Entomology,2008,53:209-230.
(责任编辑: 杨明丽)
猜你喜欢
江苏农业学报(2022年3期)2022-07-16
农村百事通(2018年14期)2018-10-31
江苏农业科学(2017年1期)2017-02-27
江苏农业科学(2017年1期)2017-02-27
湖北农业科学(2016年20期)2017-02-15
西北农林科技大学学报(社会科学版)(2017年1期)2017-02-10
科学与财富(2016年18期)2016-12-22
