
青海玛多梭罗以礼草生存群落种间关联及生态位
2019-12-12 07:31才文代吉索南才仁温小成王生彩杨毛吉仁增欧珠孙海群
草业科学 2019年11期
张 静,才文代吉,索南才仁,温小成,王生彩,杨毛吉,仁增欧珠,孙海群
(青海大学农牧学院,西宁 青海 810016)
物种的种间关联和生态位都是解释群落中物种共存与竞争机制的基本理论,是生态学的研究热点之一[1]。种间关联是不同物种在空间分布出现的相似性,体现了群落在一定时期内部物种间相互作用和影响的有机联系,是对一定时期内植物群落物种关系的静态描述[2-3]。根据物种相互间的吸引或排斥及对生境的选择和要求上,种间相关可分为正相关、负相关和不相关3种类型。我国关于物种间相关性的研究开始于70年代末[4-6],研究范围涉及森林[7]、湿地[8]、荒漠[9]、草地[10]等群落,缺少对高寒地区植物群落的种间相关性研究。生态位的理论是Grinnell于1917年首次提出[11],Hutchinson于1957年从空间和环境资源利用等方面提出“n维超体积生态位”,奠定了现代生态位理论的基础。20世纪70年代后开始应用到植物种群的研究中[12-13]。生态位是物种在特定生态环境下,物种与环境二者之间相互作用的特征表现[14],以生态位宽度和生态位重叠作为其测度指标,用于量化种群对资源利用能力及在生态系统中的功能和地位[15]。国内外对生态位的研究多集中于空间生态位上[16],关于n维梯度上的生态位研究较少,且研究的资源维度主要涉及土壤有机质、盐分、pH、含水量等因子梯度[17-19]。对种群的种间关联性和生态位研究有利于了解物种在群落中对资源的利用程度、物种之间共存与竞争的关系,可提高对种群的相互关系、群落的组成、功能及演替规律的正确认识,同时可揭示物种与环境之间的耦合关系[20-22]。
分析重点保护植物在群落中的种间关系和生态地位是保护珍稀濒危植物的重要基础,近年来,很多学者对此进行了深入的研究。许金石等[23]通过对子午岭地区内19种草本药用植物两两之间及优势木本植物之间的种间联结性的研究,提出可在子午岭地区森林生长较好的地方种植玉竹(Polygonatum odaoatum)、天门冬(Asparagus cochinchinensis)和牛蒡(Arctium lappa)等药用植物。张孝然等[24]对大黄花虾脊兰(Calanthe sieboldii)的生态位进行分析,发现在其生存的小群落内,大黄花虾脊兰对资源利用的能力相对较强,具有明显的优势地位。俞筱押等[25]通过对贵州茂兰的四药门花(Loropetalum subcordatum)群落的乔木层、灌木层和草本层的优势种群进行生态位特征分析,发现导致四药门花濒危的重要原因之一可能是与常绿物种之间存在的潜在竞争关系。曾洪和陈小红[26]在研究圆叶玉兰(Oyama sinensis)的生态位时,发现虽然圆叶玉兰具有较强的生态适应能力,但是面临着地理分布狭窄和数量较少的问题,提出要加强生境的保护及生殖繁殖方面的研究。
梭罗以礼草(Kengyilia thoroldiana)隶属禾本科以礼草属,是青海省二级保护的多年生草本植物,是亚洲中部高海拔(3 500-5 000 m)高寒草原的特有种,具有极强的抗旱抗寒能力,适应寒冷干旱的高寒草原和高寒荒漠生境,为青海高原地区该属植物的绝对优势种群[27-29]。作为高寒草原的生态草种,梭罗以礼草常被用作对高寒地区退化草地植被恢复的优质牧草,该功能对三江源区及整个青藏高原的环境保护、草地植被和生态环境恢复均具有重要意义[30-32]。
为了进一步了解梭罗以礼草的生态地位,本研究对梭罗以礼草及其周围物种之间的种间关联和多维生态位进行研究,了解梭罗以礼草所处的自然生境状态和与周围植物在环境资源因子上的相互关系,以此阐明梭罗以礼草在天然草地的生长地位,为梭罗以礼草的科学保护提供理论和实践依据。
1 研究区域与研究方法
1.1 研究地概况
玛多县位于青海省南部,是黄河发源地,境内河流密集、湖泊众多,全县共有大小湖泊4 077个,素有“千湖之县”、“黄河之源”的美誉。20世纪70年代的玛多县水草丰美,牛羊成群,发达的畜牧业使之一度成为全国闻名的富裕县。然而,随着人类不断对草原的践踏和全球气候的异变,玛多县的生态系统遭到严重的摧残和破坏,加剧草原退化、沙化,加上鼠害猖獗,导致草原水土流失严重,使之成为三江源区生态系统退化最 为 严 重 的 省 份[33-35]。研 究 地(97°43′29.52″ E,34°45′56.83″ N)位于玛多县玛查里镇,海拔4 329 m,属高平原地区,地形为起伏不大的平滩地。该地气候为典型的高原大陆性气候,一年之中无四季之分,只有冷暖之别,年平均气温-4.1 ℃,除5月-9月,各月平均气温在-3.0 ℃以下,最冷的1月均温为-16.8 ℃,极端日最低温-48.1 ℃,是青海省极端日气温最低的地方;最热月7月均温为7.5 ℃,极端日最高温22.9 ℃,温差大,全年无绝对无霜期。年均降水量303.9 mm[36-37]。该研究地植被类型为紫花针茅(Stipa purpurea)高寒草原,植被总盖度为50 %,优势种为紫花针茅,亚优势种为早熟禾(Poa annua)和梭罗以礼草,伴生种有细叶亚菊(Ajania tenuifolia)、垂穗披碱草(Elymus nutans)、长茎藁本(Ligusticum thomsonii)、粗壮嵩草(Kobresia robusta)、铁 棒 锤(Aconitum pendulum)、黄 花 棘 豆(Oxytropis ochrocephala)、麦 瓶 草(Silene conoidea)、甘肃马先蒿(Pedicularis kansuensis)、葶苈(Draba nemorosa)等。
1.2 研究方法
1.2.1 数据采集
于2018年7月的植物生长茂盛时期,基于野外勘查,在青海省玛多县玛查理镇选取一个梭罗以礼草生长旺盛的典型群落。根据梭罗以礼草现有分布及研究地地形的实际情况,由南到北等间距(50 m)平行设置5条宽2 m的样带,于每条样带上等间距(50 m)设置3个2 m × 2 m的小样地,每个小样地随机布设1个0.5 m × 0.5 m的样方,并用GPS定位,共计15个样方。记录样方内每种植物的名称、高度和盖度,同时平齐地面剪取其地上部分称量鲜重。在每个样方的4个角及中心位置采用土钻法采集0-30 cm土层的土壤,混匀,装入自封袋,作为该样方的土样。将土样带回实验室,采用烘干法测土壤含水量,重铬酸钾氧化外加热法测量土壤有机质(SOM),碱解扩散吸收法测量土壤碱解氮(AN),碳酸氢钠浸提-钼锑抗比色法测量土壤速效磷(AP),醋酸铵浸提火焰光度法测量土壤速效钾(AK),电位法测定土壤pH[38-39]。
1.2.2 数据处理
整个样地共记录21种植物,为了更好地比较梭罗以礼草种群与其他种群的种间关系,根据各个物种重要值的大小及出现的频度(剔除频度 < 7%,频度以物种出现的样方数目占总样方数的百分比进行测算)[40],选取12个常见物种(表1),形成12 × 15的数据矩阵,比较梭罗以礼草种群与样地中常见植物种群间的关系。

表 1 群落中12个常见物种编号种名及序号Table 1 Species and number of 12 common plant species in the community
1)物种重要值(P)的计算:

2)总体关联性分析:
采用方差比率(VR)法检验12个常见物种的总体关联性,采用统计量(W)判断关联性是否显著[2]。
3)种对间联结性分析:
根据物种在样方中存在与否将12 × 15的数据矩阵转化为(0,1)形式的二元数据矩阵,使用Yates连续校正的χ2公式进行χ2检验[2,5,7]。χ2检验只能定性的确定种对间的关联性质,不能定量评价关联程度,故采用Ochiai指数来评价[41-42]。
4)种对间相关性测定:
为了对样地数据进行深入的对比,参照张金屯[2]的方法,采用物种重要值指标处理数据,进行Pearson积矩相关系数和Spearman秩相关系数检验。
5)划分资源等级梯度:
依据试验测得土壤碱解氮(AN)、速效磷(AP)、速效钾(AK)、pH值、SOM和含水量的取值范围,将6个土壤因子等距划分为7个资源等级梯度[43](表2)。根据12 × 15的数据矩阵和表2,计算物种在每个土壤因子梯度下的重要值。
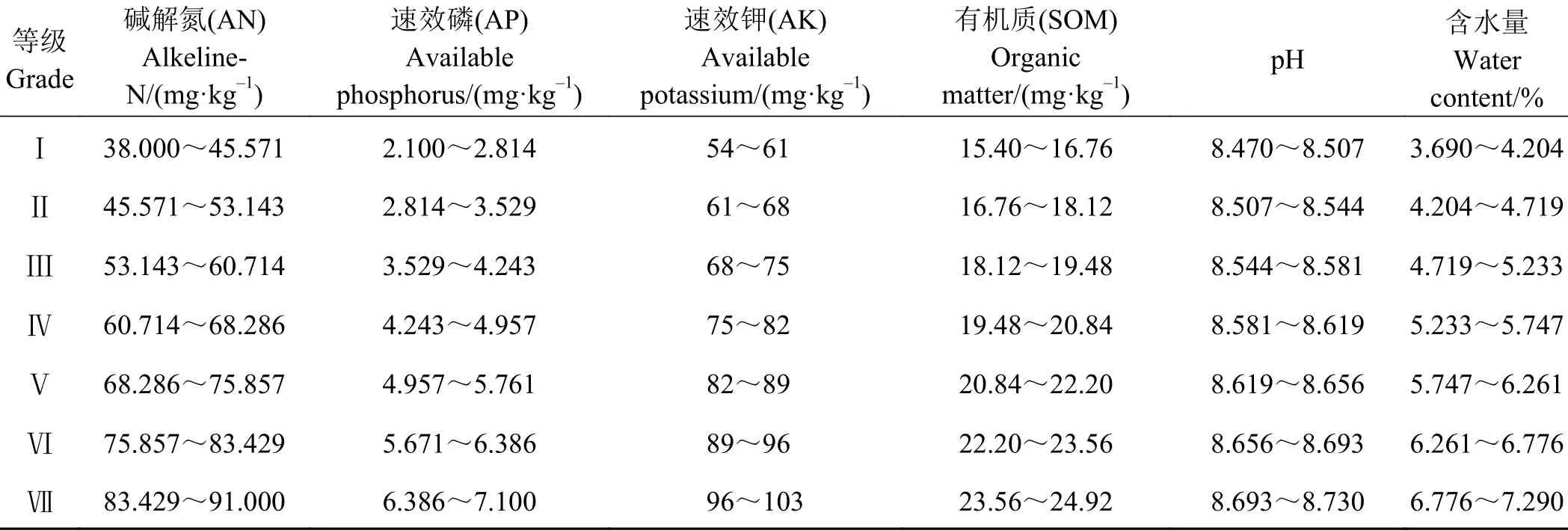
表 2 6个土壤因子等级梯度划分Table 2 Resource hierarchy of six soil factors
6)生态位宽度与生态位重叠指数计算:
采用经Corwdl修正的Levins公式(1968)测定生态位宽度[44-45];采用Pianka生态位重叠公式[19]测定生态位重叠。
样地中所有数据均使用Excel 2010及SPSS 25.00软件进行统计分析。使用SPSS 25.00计算相关系数;使用Excel 2010计算物种重要值、多物种总体关联性、χ2检验、关联指数、划分资源等级梯度、生态位宽度指数和生态位重叠指数并绘制相应图表。
2 结果与分析
2.1 多物种总体联结性
经计算方差比率(VR)测定值为0.778,小于1 (表3)。否定零假设,接受被择假设,表明种间总体关联性呈净的负关联,以自由度查相应χ2值,χ20.95(15) =7.26,χ20.05(15) = 25.00,检 验 统 计 量W = 11.677,刚好落在二者之间,表明群落多物种总体关联性不明显,反映出梭罗以礼草所在群落物种种间总体间虽然存在关联性,但是关联性不显著,物种仍趋于独立分布。

表 3 种间总体关联性检验结果Table 3 Macro-associations among the populations of the 12 studied species
2.2 种间关联
表4为Yates公式纠正后的χ2检验和Ochiai指数半矩阵表。χ2检验结果表明,在12个物种组成的66个种对中,无显著和极显著关联种对出现,说明该群落中的主要种群之间的联结性较弱,关系松散,各自趋向于独立分布。有34个种对存在正相关,32个种对存在负相关关系,正负相关比为1.06。梭罗以礼草与8种植物正关联,其中与紫花针茅、早熟禾、细叶亚菊的联结值最大,表现为较强的正联结关系,与另外3种植物呈负关联关系,联结强弱依次为盐地风毛菊(Saussurea salsa) > 长茎藁本 > 垂穗披碱草。
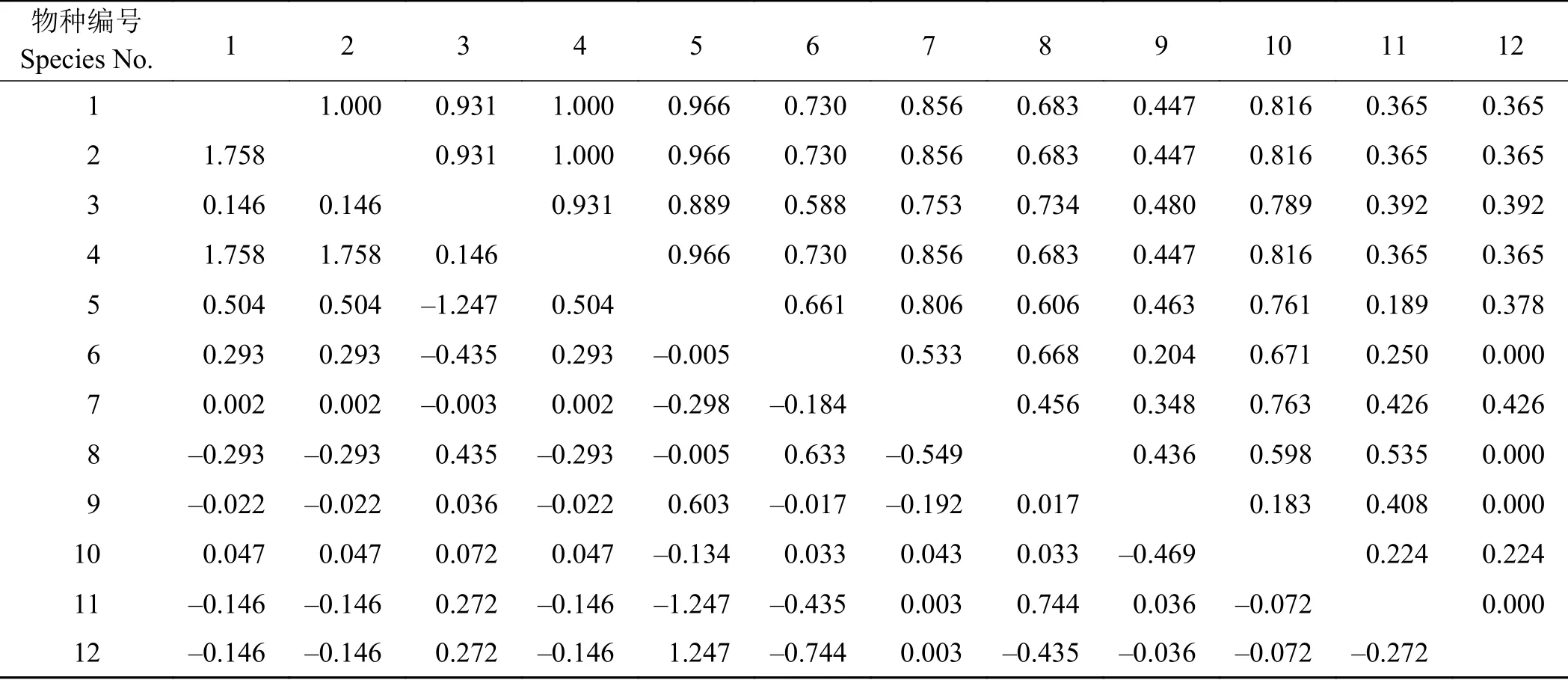
表 4 物种间 χ2检验及Ochiai指数半矩阵表Table 4 Semi-matrix table of interspecies associations, with χ2-test values and Ochiai indices values for the 12 studied species
由于紫花针茅、早熟禾和细叶亚菊3个物种出现在所有样方中,它们与其他物种的关系仅有两种情况:①两个物种都存在;②紫花针茅、早熟禾或细叶亚菊存在而其他种不存在。频度达到100%的3个物种,使得b和d均为0,因而无法进行χ2检验。但这并不表明紫花针茅、早熟禾和细叶亚菊与其他物种之间不存在关联,本研究中采用王伯荪提出的给b和d值加权为1进行计算,结果表明,紫花针茅与其他物种间不存在显著关联。早熟禾和细叶亚菊的结果同上。
Ochiai指数是用来描述种对间的关联程度,将该指数值分为6个层次,根据层次内种对数由大到小依次为0.6~0.8、0.8~1、0.4~0.6、1、0.2~0.4、0~0.2。指数结果表明,物种对在0.6~0.8中的种对数占很大优势,而0~0.2的指数值仅占极少数,表明大部分种对间正(负)关联程度居中。此外,紫花针茅-早熟禾、紫花针茅-细叶亚菊和早熟禾-细叶亚菊3个种对间存在极显著关联。梭罗以礼草与其他植物的指数值范围主要集中在0.8~1且没有特别不相关的种对出现,与之连接最紧密的为紫花针茅、早熟禾和细叶亚菊,指数值均在0.9以上。
2.3 相关分析
Pearson积矩相关系数和Spearman秩相关系数是反映两个物种种间协变线性关系的重要指标,可反映物种间的数量变化关系[9]。Pearson积矩相关系数检验(表5)表明,梭罗以礼草所在群落内的12种物种种群间,有正相关关系和负相关关系的均有33对,其中,显著正相关、显著负相关和极显著负相关各有一对,分别为粗壮嵩草-阿尔泰草(Koeleria macrantha)、紫花针茅-盐地风毛菊和早熟禾-火绒草(Leontopodium leontopodioides)。Spearman秩相关系数检验(表5)表明,66个种对中,有32对属于正相关关系,33对属于负相关关系,其中,有3对显著正相关(早熟禾-细叶亚菊、粗壮嵩草-火绒草、粗壮嵩草-阿尔泰草)、1对极显著负相关(细叶亚菊-阿尔泰草)和4对显著负相关(早熟禾-垂穗披碱草、细叶亚菊-粗壮嵩草、早熟禾-火绒草和盐生风毛菊-萎软紫菀(Aster flaccidus)。
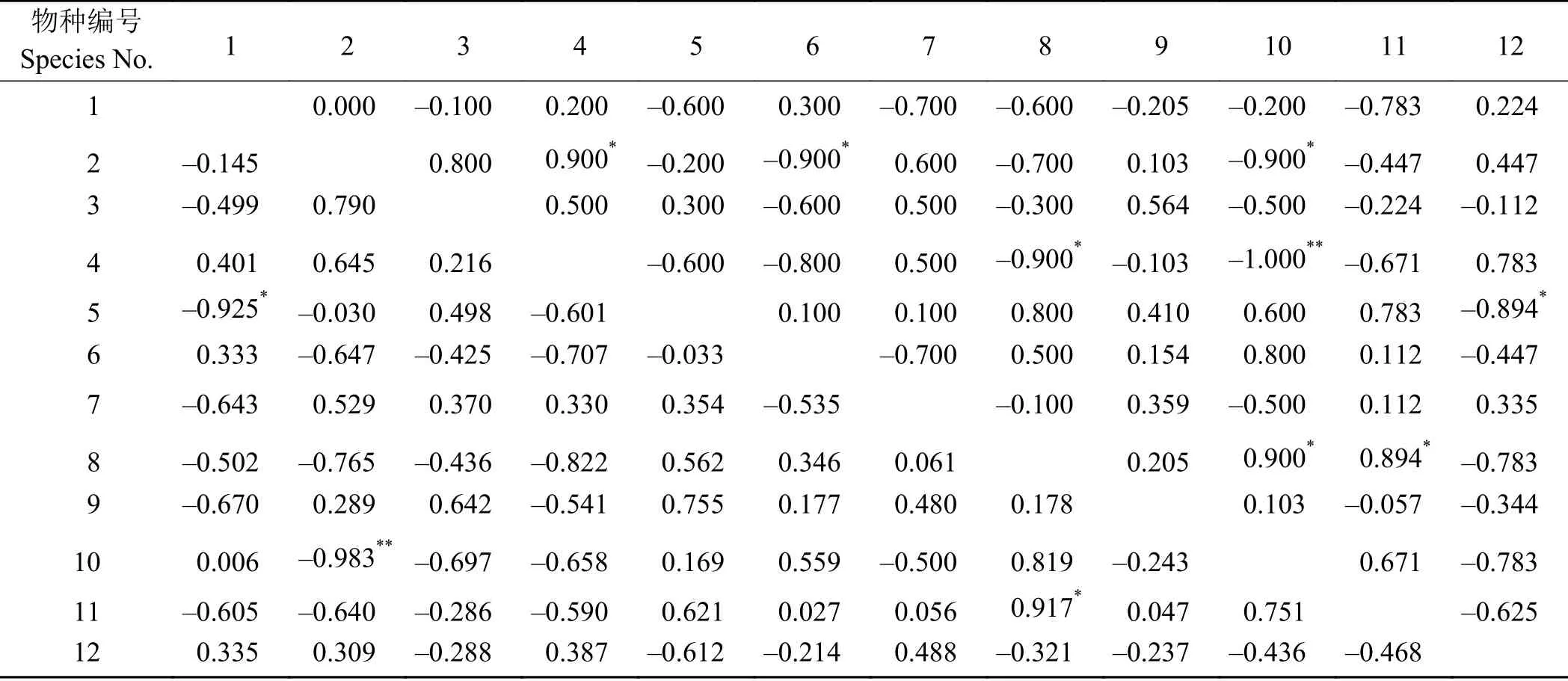
表 5 物种间Pearson积矩相关系数和Spearman秩相关系数半矩阵表Table 5 Semi-matrix table of Pearson's correlation coefficients and Spearman's rank correlation coefficients for the 12 studied species
通过对Pearson积矩相关系数和Spearman秩相关系数的计算,结果具有相似的变化(表6)。二者均表明,12个物种间呈现显著或极显著关联的种对相对较少,且梭罗以礼草与其他11种植物间没有出现显著或极显著的关联,无显著关联种对和极显著关联种对的出现,说明该群落中的主要种群之间的联结性较弱,关系松散,各自趋向于独立分布,这点与χ2检验的结果一致。梭罗以礼草与5种植物有正关联,关联强度较强为早熟禾和黄花蒿(Artemisia annua);与6种植物有负关联,关联强度较强为火绒草和紫花针茅。

表 6 χ2检验、Pearson积矩相关系数、Spearman秩相关系数检验结果Table 6 χ2-test results, Pearson's correlation coefficients, and Spearman's rank correlation coefficients
2.4 生态位宽度
Levins (1968) 生态位宽度指数可以用来描述物种利用资源的能力总和。根据对12个主要物种在6个土壤因子梯度水平上的统计计算(表7),发现物种的生态位宽度与土壤因子梯度间存在复杂的关系,主要表现在,同一物种在不同土壤因子梯度上的生态位宽度不同,如频度为100 %的紫花针茅、早熟禾和细叶亚菊在AN因子梯度上的生态位宽度分别为6.604、6.814和6.133,而在含水量因子梯度上生态位宽度分别为5.881、5.746和4.912,在AP因子梯度上生态位宽度分别为4.404、4.745和4.838。同一土壤因子梯度上的不同物种的生态位宽度不同,在6个因子梯度上均表现较宽的物种大多为该群落的优势种。在AK、SOM和含水量因子梯度上,生态位宽度较大的物种为紫花针茅、早熟禾、梭罗以礼草;在AN和pH值因子梯度上,生态位宽度较大的物种为紫花针茅、早熟禾、盐地风毛菊;在AP因子梯度上,生态位宽度较大的物种为梭罗以礼草、长茎藁本、粗壮嵩草。12个常见物种在6个因子梯度上的生态位宽度均值大小顺序依次为:紫花针茅 > 早熟禾 > 梭罗以礼草 > 盐地风毛菊 > 长茎藁本 > 细叶亚菊 >粗壮嵩草 > 火绒草 > 垂穗披碱草 > 黄花蒿 > 阿尔泰草 > 萎软紫菀;12个常见物种在每个因子梯度上的生态位宽度均值排列顺序为SOM > pH > AN >含水量 > AK > AP。

表 7 6个土壤因子梯度上12个常见物种生态位宽度Table 7 Niche breadths of the 12 studied species across six soil factor gradients
在AN水平上,早熟禾的生态位宽度值均位居首位,梭罗以礼草处于第6位,是在6个水平中最窄的;在AP和SOM水平上,梭罗以礼草的生态位宽度值都是最大的;在AK水平上,紫花针茅的生态位宽度值都位居首位,梭罗以礼草处于第3位;在pH水平上,紫花针茅的生态位宽度值均位居首位,梭罗以礼草处于第4位;在含水量水平上,紫花针茅的生态位宽度值都位居首位,梭罗以礼草的均位于其次。通过总均值的计算发现,样地中生态位宽度值最大的是紫花针茅,其次为早熟禾,梭罗以礼草处于第3位。总之,在本样地中梭罗以礼草种群表现出了较宽的生态位,特别是在AP和SOM水平上,可充分利用土壤AP和SOM。
2.5 生态位重叠
12种植物组成的66个种对在6个土壤因子梯度上生态位重叠值如表8、9、10所列。总体上看,本研究区的12种植物的生态位重叠值较高。为了更好地表现12个物种的生态位重叠情况,将生态位重叠值划分为以下3个类型:重叠值 >0.9,为高重叠;0.5 < 重叠值 < 0.9,为中重叠;重叠值 ≤ 0.5,为低重叠。在AN因子梯度上,属于高重叠的有9对,占总对数的13.64%;属于中重叠的有42对,占总对数的63.64%;属于低重叠的有15对,占总对数的22.73%。在AP因子梯度上,属于高重叠的有22对,占总对数的33.33%;属于中重叠的38对,占总对数的57.58%;属于低重叠的仅有6对,占总对数的9.09%。在AK因子梯度上,属于高重叠的有7对,占总对数的10.61%;属于中重叠的有39对,占总对数的59.09%;属于低重叠的有20对,占总对数的30.30%。在SOM因子梯度上,属于高重叠的有5对,占总对数的7.58%;属于中重叠的有46对,占总对数的69.70 %;属于低重叠的有15对,占总对数的22.73%。在pH因子梯度上,属于高重叠的有8对,占总对数的12.12 %;属于中重叠的有40对,占总对数的60.61%;属于低重叠的有18对,占总对数的27.27%。在含水量因子梯度上,属于高重叠的有16对,占总对数的24.24%;属于中重叠的有38对,占总对数的57.58%;属于低重叠的有12对,占总对数的18.18%。由此可见,12种植物间的生态位重叠值偏高,对资源利用的相似程度较高,种间可能存在较高的竞争关系。

表 8 12种常见植物66个物种对在速效磷和碱解氮因子梯度上生态位重叠Table 8 Niche overlaps of 66 species of 12 common plants across AP and AN soil factor gradients

表 9 12种常见植物66个物种对在有机质和pH因子梯度上生态位重叠Table 9 Niche overlaps of 66 species of 12 common plants across SOM and pH soil factor gradients
在SOM和AK水平上,与梭罗以礼草的生态位重叠程度较大的依次为紫花针茅、早熟禾、盐地风毛菊、长茎藁本;在pH水平上,与梭罗以礼草的生态位重叠程度较大的依次为早熟禾、盐地风毛菊、粗壮嵩草、紫花针茅;在AP水平上,与梭罗以礼草的生态位重叠程度较大的依次为粗壮嵩草、紫花针茅、长茎藁本、盐地风毛菊;在AN水平上,与梭罗以礼草的生态位重叠程度较大的依次为早熟禾、粗壮嵩草、盐地风毛菊、紫花针茅;在含水量水平上,与梭罗以礼草的生态位重叠程度较大的依次为盐地风毛菊、紫花针茅、早熟禾、火绒草。其中,与梭罗以礼草生态位重叠值最大的是在含水量水平上的盐地风毛菊,二者的重叠值达到0.988,表明二者在争夺水分时可能存在较强的竞争关系。整体上看,梭罗以礼草与其他11种植物在6个资源水平上共组成66个种对,其中生态位重叠值大于0.9的有22对,占总对数的33.33%,表明了梭罗以礼草种群与群落内其他物种种群在资源利用方式上极其相似,同时也预示了它们之间极易发生激烈的竞争。

表 10 12种常见植物66个物种对在含水量和速效钾因子梯度上生态位重叠Table 10 Niche overlaps of 66 species of 12 common plants across the water level and AK soil factor gradients
3 讨论
3.1 种间关联
χ2检验、Pearson积矩相关系数和Spearman秩相关系数的检验结果存在一定的差异(表4、表5、表6),如粗壮嵩草-阿尔泰草种对在χ2检验时仅呈现正关联,而在相关检验中呈现出显著性。这主要是因为在进行χ2检验时物种丧失了种间相关的多度信息,而相关分析中应用了物种重要值,所以做定量分析的相关系数检验比做定性判断的χ2检验更准确。此外Pearson积矩相关系数是以样本为正态分布为计算前提,但自然生境下的物种多为聚集分布,所以非参数检验的Spearman秩相关系数检验体现了较高的灵敏度且更符合实际[46]。
许多研究表明[47-48],在一般条件下,群落处于演替初期时,物种的种间联结程度往往处于较低水平,主要表现为负关联,且物种间存在激烈竞争;随着群落演替的不断发展,群落的结构及种类组成将逐渐趋向于不断完善和稳定,物种种间正关联的程度呈现出不断增大的现象,以求达到多物种间的稳定共存状态。在本研究的12个常见植物中,大多数物种对生境的要求比较低,不仅生长于紫花针茅高寒草原,也常见于高山嵩草草原化草甸等草地类型,如细叶亚菊和早熟禾常作为伴生种出现在高山嵩草+细柄茅草地型及赖草型等草地类型中。这可能导致了66个种对在χ2检验和相关系数检验中正负关联比例十分接近且显著和极显著的种对相对较少(表6),物种间关联性较弱,各自更趋向于独立分布。同时,考虑到本样地的植被覆盖度只有50 %,可见研究群落内种群的分布未超出环境容纳量,群落很有可能正处于植被演替进程中一个相对稳定的过渡阶段。
3.2 生态位
生态位宽度能够衡量物种对环境资源利用状况的尺度,反映物种在栖息地中的地位和分布状况,在一定程度上,数值越高表示物种对资源的利用程度越强,分布越均匀,在群落中往往处于优势地位[2]。在该样地中,梭罗以礼草的重要值相对较高且在6个土壤因子梯度上同时表现出了较大的生态位宽度,极强的抗旱抗寒能力使得梭罗以礼草对这6种土壤因子的利用能力和对环境的适应能力显著高于其他很多物种,更能充分利用对土壤AP和SOM,分布范围较广,有可能处在群落中的优势地位[49],对维持群落内部环境以及生态环境的稳定起着重要作用。此结论与一些生长于特殊生境中的重点保护植物具有较大的生态位宽度的研究结果一致[26]。
生态位重叠反映了植物对同等级资源的利用程度以及空间配置关系,表明物种之间对资源利用的相似程度和竞争关系[50]。一般认为,种间竞争越激烈,生态位重叠越大,实际上生态位重叠并不一定能引起竞争,资源丰富时生态位重叠值高,只表明两个物种因生态相似性而占据了相近的生态空间[26]。因此,生态位重叠程度较高的两种植物种群生活在同一生境,对相同资源进行竞争时,它们之间存在两个可能的关系,一是由于资源共享进行竞争,此时双方具有显著的正、负关联;二是在空间、资源充足的条件下,由于利用资源的相似性彼此共存,此时双方种间关联不显著[47,49]。整体上看,梭罗以礼草与周围植物在土壤因子梯度上的生态位重叠值显著高于其他种对。由此可见,为更好适应高海拔气候及维持种群繁衍,梭罗以礼草种群要尽可能汲取较多的养分,在这个过程中,可能遇到同样为生存汲取养分的其他植物,加上研究地恶劣的生境条件,物种间很容易出现激烈竞争。其中,在含水量因子梯度上,盐地风毛菊与梭罗以礼草有高重叠值和高宽度值且十分相近,说明二者对土壤水资源的利用方式很相似,很可能与二者的生长习性有关。盐地风毛菊多生于戈壁滩和湖边等地,梭罗以礼草多生于山坡草地和河岸坡地等地。同时二者的种间关系存在一定的正相关,表明二者在土壤水份因子上很可能是互利共生的。调查显示,自然生境中梭罗以礼草的平均植株高度约为17.371 cm,显著高于周围植物,无伴生的木本植物,这些特征表现出它的耐旱性和耐寒性,是适应寒冷干旱的高寒草原和高寒荒漠的环境,与其他种群形成共存关系的基础。
有研究表明,生态位宽度值较窄的物种之间生态位重叠值较高;生态位宽度值较宽的物种之间生态位重叠值较低[9]。本研究发现,垂穗披碱草与火绒草在AK水平上生态位宽度值都较小但却有较高重叠值,说明生态位宽度值较小的物种之间有可能出现较高的生态位重叠。盐地风毛菊与紫花针茅在SOM水平上生态位宽度值都较大,生态位重叠为高重叠,说明生态位宽度值较大的物种之间有可能出现较高的生态位重叠。故生态位宽度与生态位重叠之间并不存在直接的线性关系[9]。
3.3 种间关联与生态位的关系
植物种群间的生态位重叠和种间关联性都是由资源竞争的重叠性而引起,二者皆为表征种间相互作用的重要因素,因此存在着一定的关联性且相关性较强。很多研究表明,正关联显著的种对,一般有高的生态位重叠值;而负关联显著的种对,重叠值很小或为零[51]。然而,这些结果建立在对物种间生态位的单维水平上,并未直接考虑深入研究物种在多种环境因子的作用。本研究发现,正关联较高的种对,在6个维度上均表现出了相对较大的生态位重叠,如早熟禾和细叶亚菊在AN、AP、AK、SOM、pH和含水量因子梯度上的生态位重叠值依次为0.945、0.728、0.885、0.870、0.871和0.912。负关联较高的种对,仅在某几个生态位维度上表现出了相对较大的生态位重叠,整体上重叠程度不如正联结较高的种对,如细叶亚菊和粗壮嵩草在AN、AP含水量因子梯度上的生态位重叠值依次为0.826、0.883和0.793,而在AK和SOM因子梯度上的生态位重叠值分别为0.355和0.463。此结果与陈旭等的观点存在差异,陈旭等[52]认为种间的正关联性越强,不一定说明所有维度上生态位重叠值就越大,仅在某一个或几个生态位维度上表现出较大的重叠;而由资源竞争引起的负关联生态位重叠值高,由生境差异导致的负关联生态位重叠值低。首先,二者的研究对象不同,陈旭等[52]研究的是长白山哈泥泥炭地的苔藓植物,本研究中是玛查理镇的高原植物,两类物种本身特性及不同的生境有可能对最后的结果造成间接影响。另一方面,二者研究的维度种类及个数存在差异。陈旭等[52]除选取pH等土壤因子梯度外,还涉及到乔木郁闭度等环境因子梯度,从9个维度上较为全面分析了7种苔藓植物的生态位。而本研究只选取了含水量等6个土壤因子梯度,所涉及到的环境因子梯度较少,这可能造成研究的物种在其他环境因子梯度上产生分异,从而直接影响到研究结果。
此外,由于种群之间关联性和生态位的研究受时间的限制,随着群落的演替发展,植物群落的自然分布与气候因子、立地条件和调查时间关系密切,在种群发育的时期,植物的高度和盖度等会发生变化。本文仅对梭罗以礼草目前生存现状进行研究;今后还需对梭罗以礼草的种群动态、生殖生理方面做进一步的探究,为保护濒危植物梭罗以礼草提供更多的科学依据。
4 结论
本研究采用种群种间关联和生态位相结合的方法对自然植被下梭罗以礼草所分布的群落中常见种群进行分析,结果表明,1)该群落结构简单,物种丰富度较小,群落可能正处于植被演替进程中一个相对稳定的过渡阶段。12种常见植物的物种间联结性松散,呈现相对独立分布的格局,易受外界干扰。2)梭罗以礼草与其他常见植物均无出现显著关系,彼此独立分布;对环境的适应能力和6种土壤因子尤其是土壤AP和SOM的利用能力显著高于其他很多物种,有可能处于群落中的优势地位;在对土壤因子的利用方式上,与内其他常见植物极其相似,可以相互补充,尤其表现在与盐地风毛菊在水分利用的方式上;较强生态适应能力使之成为高寒草原的生态草种和植被恢复的优质牧草。3)在12个常见植物组成的66个种对中,正关联较高的种对,在6个土壤因子梯度上均表现出了相对较大的生态位重叠;负关联较高的种对,仅在某几个土壤因子梯度上表现出了相对较大的生态位重叠,整体上重叠程度不如正联结较高的种对。
猜你喜欢
河南师范大学学报(自然科学版)(2022年5期)2022-08-08
数学物理学报(2022年1期)2022-03-16
数学物理学报(2021年6期)2021-12-21
数学物理学报(2021年5期)2021-11-19
作文评点报·作文素材初中版(2020年22期)2020-06-21
中外文摘(2019年10期)2019-11-12
华东师范大学学报(自然科学版)(2019年3期)2019-06-24
蔬菜(2018年5期)2018-05-17
新少年(2017年11期)2017-12-07
杂草学报(2015年2期)2016-01-04
