
23份豌豆材料过氧化物酶同工酶研究
2019-12-19 01:51王翠玲杨晓明王旺田周卫东
生物学杂志 2019年6期
王翠玲, 杨晓明, 王旺田, 周卫东
(1. 甘肃农业大学生命科学技术学院 甘肃省干旱生境作物学重点实验室, 兰州 730070;2. 甘肃省农业科学院粮食作物研究所, 兰州 730070, 3. 兰州现代职业学院 农林分院, 兰州 730070)
豌豆(PisumsativumL.)属豆科,蝶型花亚科,一年生或越年生攀援性草本植物,兼有蔬菜、粮食、饲料功能,是世界第二大食用豆类作物,豌豆适应能力很强,在全世界范围内有广泛的地理分布,多样化的生长条件形成了豌豆的遗传多样性[1]。豌豆还具有良好的固氮性能,能够改善土壤氮肥供应,在草地农业生态系统中发挥着非常重要的作用[2-3]。世界各地区都十分重视对豌豆种质资源的研究,对其种质资源进行收集、利用和保存[4]。中国地域广阔,豌豆资源比较丰富,其收集和研究开始于20世纪60年代,目前国家种质资源库中已保存豌豆资源5000余份[5]。豌豆种质资源的研究,可以丰富我国豌豆遗传资源基因库,提高豌豆抗逆性、抗病性研究,加快豌豆新品种培育的步伐[1,4,6]。
过氧化物酶(peroxidase, POD)是植物体内普遍存在的、活性较高的一种酶,它与呼吸作用、光合作用及生长素的氧化等都有关系。过氧化物酶同工酶酶谱的差异能够反映基因所处的遗传背景差异[7-9]。宋志文等[10]利用过氧化物酶同工酶研究中国东北地区野豌豆属植物;张建全等[11]对4个春箭舌豌豆品系原种及第3代过氧化物酶同工酶分析,探讨F3代农艺性状等发生变化的遗传机制。本研究拟对23个豌豆种质资源的过氧化物同工酶表达进行分析,结合豌豆种子形态特征,为豌豆的遗传多样性分析以及新品种选育提供科学依据。
1 材料与方法
1.1 材料
由甘肃省农科院随机抽取国内外不同地理来源豌豆材料23份。中国品种资源20份,分别来自西藏(3份)、甘肃(2份)、陕西(5份)、四川(4份)、内蒙古 (2份)、云南(1份)、台湾(1份)、宁夏 (2份);国外品种资源3份,来自澳大利亚(3份)。各材料编号特征见表1。
1.2 方法
1.2.1 种子萌发培养及酶液的提取
23份豌豆材料各选取成熟饱满的种子8~10粒浸泡,萌发,每天早晚各换MS营养液2次,14 d后,取各豌豆的根0.5 g洗净,于预冷的研钵中,加入样品提取液5 mL,研成匀浆,16 000 r/min离心10 min,取上清液,即酶液,备用。
1.2.2 酶的聚丙烯酰胺凝胶电泳[12]
采用聚丙烯酰胺垂直平板电泳。浓缩胶T=3.5%,分离胶T=7.5%,电极缓冲液pH 8.3,将酶液与样品处理液1∶1混合,每空加样15 μL,起始电压110 V,分离胶电压200 V,电泳至溴酚蓝下沿1 cm左右处停止。
1.2.3 过氧化物同工酶酶染色
电泳结束后,剥胶,采用醋酸联苯胺法进行染色,具体步骤为:将蒸馏水漂洗过的凝胶放入POD染色液中,震荡5 min待酶带显出后,用蒸馏水漂洗,拍照绘图分析。
1.2.4 数据分析
依据电泳照片,记录数据,采用美国UVP凝胶成像分析系统求出相对迁移率,计算相似系数和不相似系数。酶带的相对迁移率(Rf)=酶带的迁移距离/溴酚蓝的迁移距离;过氧化物酶同工酶酶谱间相似系数c=2w/(a+b),其中c为酶谱间相似系数,w为A和B两个种相同的酶带数,a为种A在酶谱中的酶带数,b为种B在酶谱中的酶带数。采用SPSS23.0软件对 23份豌豆材料的亲缘关系及遗传差异进行聚类分析。
2 结果与分析
2.1 不同豌豆材料幼苗根部过氧化物酶酶谱特征
23份供试豌豆材料根部过氧化物酶同工酶酶谱和酶谱模式见图1和图2。从图1和图2中可以看出:各豌豆材料过氧化物同工酶在酶带数、酶量、相对迁移率等方面存在差异,有的位点上出现稳定一致的酶带,有的位点呈现很强的特异性。供试材料中共出现161条酶带,酶带类型为14种,酶带最多是8条酶带,最少是5条酶带,有8条、7条酶带的材料为多数,少数材料酶带为6条或5条,表明23个供试豌豆材料具有较丰富的遗传多样性。
根据酶谱特征,从负极到正极按相对迁移率Rf可把它们分成3个区,即A、B、C 3个区域,A区是近阴极端的慢速迁移区,Rf=0.13~0.21,有3种类型酶带,23个豌豆材料在A区共有酶带P1,Rf=0.13。B区为中速迁移区,Rf=0.37~0.51,有4种类型酶带,在B区,23号有1条酶带,其余材料有2条酶带;在B区的共有带为P4,Rf=0.37。A区与B区各品种P1、P4的染色程度不一样,说明不同材料的酶活性差异较大,酶带染色最深为1、2、6、11、12、13、15及19号材料,而8、10、14号材料的染色最浅。C区为近阳极端的快速迁移区,Rf=0.62~0.83,有7种类型酶带,不同材料的酶带数有2到4条不等,C 区的没有公共带,C区的酶带数、酶量在不同豌豆品种间有差异较大,在染色程度上表现非常明显,但也一些比较接近,如12和13号材料。

表1 23份豌豆材料编号
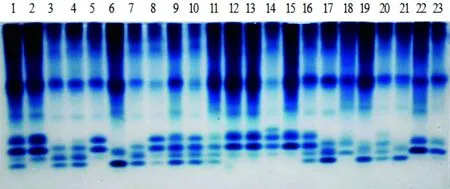
图1 23份豌豆材料豌豆幼苗根过氧化物酶同工酶电泳图谱
Figure 1 The electrophoretogram of POD isozymes in the roots of 23 cultivars ofPisumsativumL.
2.2 不同豌豆材料过氧化物酶同工酶聚类分析
亲缘关系分析聚类分析是用数学方法将一批样品(变量)按其性质的亲疏远近程度进行分类的科学方法,用酶谱间相似系数(similarity coefficient) 的大小表示分类群亲缘关系的远近,酶谱相似系数等于2个分类群中相同的酶带总数除以2个分类群中酶带总数。相似系数在0~1之间,亲缘关系越近的种类,相似系数越接近于1,彼此无关的种类,其相似系数越接近于0。利用各品种所具有的酶带数计算出相似系数c,各个种源间的遗传相似度在0.33~1.00之间,说明不同种源的豌豆种子间有不同程度的遗传差异。
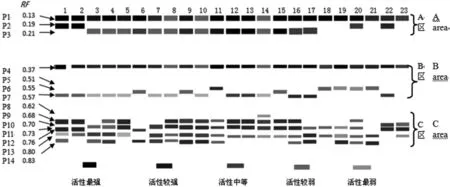
图2 23份豌豆材料根过氧化物酶同工酶酶谱模式图
采用SPSS23.0软件对23份豌豆材料的亲缘关系及遗传差异进行聚类分析,结果见图3。当取不同阈值时聚成不同类,若取0.5时,供试豌豆材料可聚为5类,分别为Ⅰ,Ⅱ,Ⅲ,Ⅳ,Ⅴ。第Ⅰ类为1、2号材料,都来源于中国西藏,全国统一编号为G0004526和G0004548,品种名都为豌豆;粒形为球形,但种皮表面有差异,1号种皮凹坑,2号光滑。第Ⅱ类为3、4、5、7、8、9、10、11、12、13、14、15、16、17号材料,共14分材料,分别来源于中国甘肃(2份)、中国陕西(5份)、中国四川(3份)、中国内蒙古 (2份)、中国云南(1份)、中国宁夏 (1份)。从品种来看,以麻豌豆居多,有 8个品种为麻豌豆,小麻豌豆、大麻豌豆、青麻豌豆、青豌豆各1份,2份豌豆。其中3号与17号材料、8号与16号材料、12号与13号材料亲缘关系相互最近,相似系数为1。第Ⅲ类为18、19、20、21、23号材料,其中18为来自中国四川的白豌豆,全国统一编号是G0005571;19号为来自中国台湾的台中11(荚豌豆);20号是来自中国宁夏的蓝豆,粒形为扁形;21和23号都来自于澳大利亚。第Ⅳ类为22号材料,自于澳大利亚的品种JI.1089,单独一类。6号材料也单独聚为一类,为第Ⅴ类,来自中国西藏的麻豌豆G0004524,粒形扁形,光滑,粒色紫黑,不同于其他材料。从聚类图来看,6号材料与其他材料的亲缘关系较远。
3 讨论
遗传多样性的表达主要是通过对遗传多态性的标记来反映的,可通过某个表型特征、特异的DNA片段或特异的同工酶带型及蛋白质作为目的基因直接或间接的标记,来研究基因的变异和遗传规律[13]。豌豆种质资源遗传多样性研究起步较晚,目前有关豌豆种质资源的遗传多样性研究主要通过相关田间农艺性状的调查来实现[14-15]。自1959年Market首次提出了同工酶的概念以来,有关同工酶的研究就很快发展起来。过氧化物酶也可以用来推测各分类群之间的亲缘关系,进行遗传多态性分析。本研究23份供试豌豆材料过氧化物酶同工酶图谱在酶带数、酶带浓度、酶带颜色深浅、扩散程度及迁移率上都有明显差别。对过氧化物酶同工酶酶谱分析,由负极向正极可分为A、B、C 3个区域,分离出161条酶带,酶带类型为14种,酶带最多是8条酶带,最少是5条酶带,A区和B区各有一条共有表达酶带,C区Rf=0.62~0.83,有7种类型酶带,没有共有酶带,但C区的酶带数、酶带颜色深浅变化丰富,表现出较强的多态性。23份材料中有两条共有酶带,又有各自的特征酶带,既显示出豌豆物种具有一定程度的遗传稳定性,也蕴涵着丰富的区域差异性及遗传多样性。
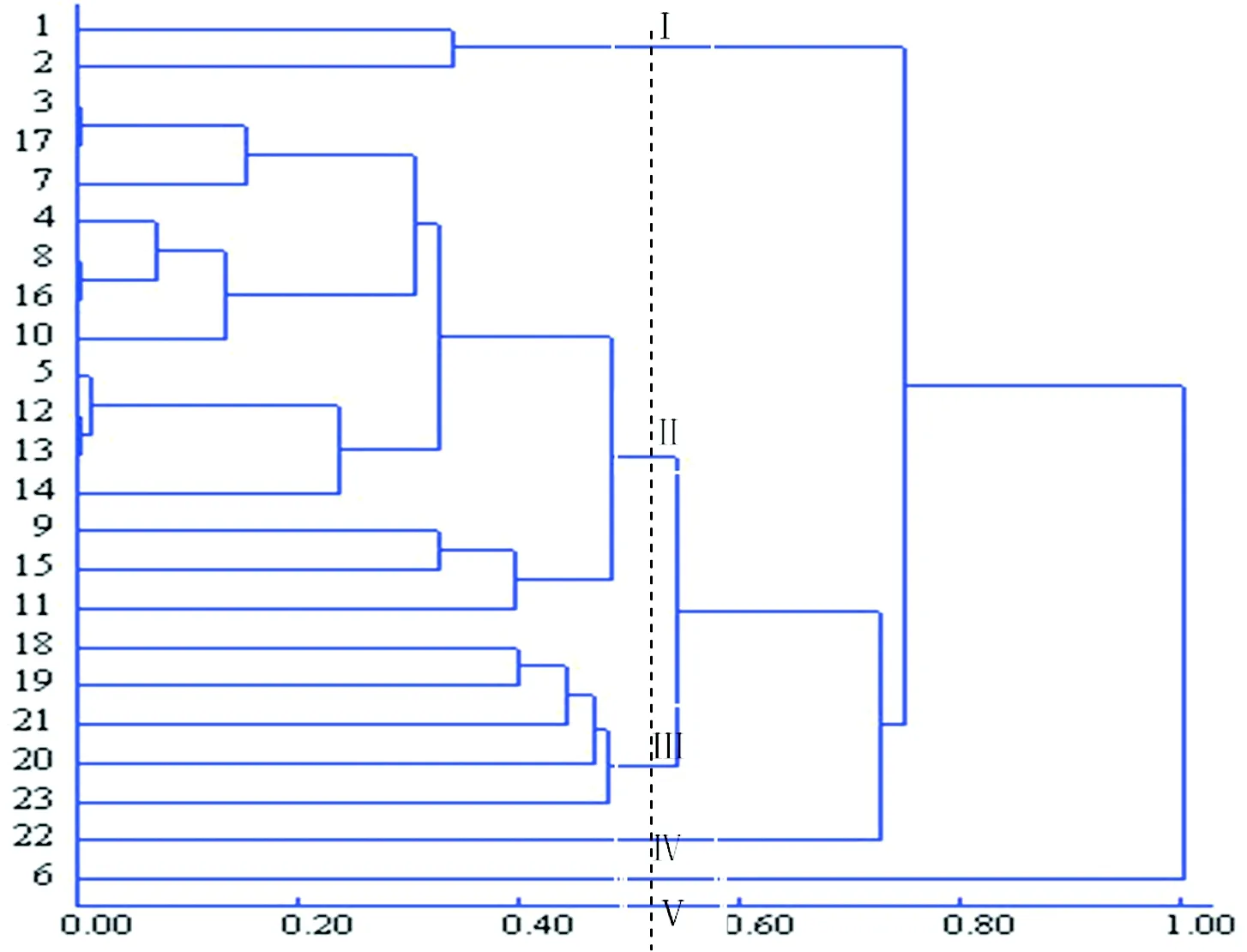
图3 23份豌豆材料同工酶酶谱性状聚类图
酶谱相似系数计算所得23份供试豌豆材料间的遗传相似度在0.33~1.00之间,聚类分析结果表明:1、2号豌豆材料聚为一类,都来自于中国西藏,比较1、2号材料的酶谱,A区、B区酶谱特征几乎一样、C区2号材料少了一条酶带。来自于中国6个省和自治区的麻豌豆系列聚为一大类,其中3号与17号材料、8号与16号材料、12号与13号材料相互间的亲缘关系最近,酶带几乎一样,相似系数为1。来自于中国四川的白豌豆,中国宁夏的蓝豆,中国台湾的台中11(荚豌豆),澳大利亚的IFPI 3917、CRIMSON ROSE聚为一大类。来自于澳大利亚的品种JI.1089,单独一类; 6号材料中国西藏麻豌豆G0004524,也单独聚为一类,其酶带数较少,为6条,与其他材料的亲缘关系最远。以上结果表明豌豆过氧化物酶同工酶与品种有关系,如大多数麻豌豆系列聚为一大类;也与其地理分布有着很大的关系。曾亮用ISSR标记技术对来自国内外的73份豌豆材料进行遗传多样性分析,基本符合地理来源相近的材料聚为一类,呈现出一定的地域性分布规律;曾亮还对120份豌豆材料的20个表型性状分析,表明豌豆属植物种质资源形态变异十分丰富,国内和国外群体内差异显著[6]。本研究中豌豆分类与豌豆品种的粒形颜色有关,如来源于中国西藏的麻豌豆,粒形扁球、光滑、紫黑,不同于其他麻豌豆,其亲缘关系与其它材料较远,单独为一类。李红等在对不同苜蓿品种过氧化物酶同工酶的分析中也得出,品种间的地理位置越近,同工酶也就越相似,酶谱的相似程度也就越高,相反则低,可以根据同工酶的差异来对品种进行分类[16]。刘贺贺等应用POD同工酶分析75份蕨麻材料的遗传多样性,结果与形态学标记、细胞水平、分子标记研究的结果一致[17]。利用过氧化物酶同工酶来探讨种质遗传关系,具有周期短、成本低、操作简便等优势。本研究的不足之处是所选用的材料较少,如果能与形态学、细胞学等研究相结合,分类会更准确。
总之,本试验的23份材料过氧化物酶同工酶酶谱分析结果表明:供试材料在同工酶系统中均表现出一些明显的共同特征谱带,显示出豌豆物种具有一定程度的遗传稳定性,同时各材料的同工酶酶谱之间也表现出某些明显的差异性,体现出豌豆物种蕴涵着丰富的区域差异性及遗传多样性,为豌豆进化、分类、种质资源的保存及利用奠定基础。
猜你喜欢
作物学报(2022年2期)2022-11-06
河北科技师范学院学报(2022年2期)2022-08-26
生命科学研究(2021年1期)2021-01-25
微生物学杂志(2020年6期)2020-03-05
天然产物研究与开发(2018年10期)2018-11-06
分析化学(2018年7期)2018-09-17
广东农业科学(2017年5期)2017-08-29
江苏农业科学(2016年8期)2017-02-15
中国塑料(2016年3期)2016-06-15
河北渔业(2015年9期)2015-09-15
