
旱涝交替胁迫对水稻荧光参数与光合特性的影响
2019-12-31 06:27俞双恩刘子鑫高世凯丁继辉
农业机械学报 2019年12期
俞双恩 刘子鑫 高世凯 王 梅 丁继辉
(1.河海大学农业工程学院, 南京 210098; 2.华北水利水电大学水利学院, 郑州 450045)
0 引言
水稻是我国重要的粮食作物之一。水资源紧缺是限制我国水稻面积发展的主要因素,不合理的灌溉制度不仅会造成水资源浪费、水稻减产,甚至会形成农业面源污染,破坏生态环境。水稻控制灌排技术具有节水、减排、高产、控污效应[1-4]。水稻控制灌排技术的要点是,在灌溉期稻田处于干湿交替的环境,雨季稻田滞蓄一定深度的水层。因此进行旱涝交替胁迫机理研究对完善控制灌排理论体系具有重要意义[5-6]。
植物90%以上的干物质累积和产量形成来自叶片光合作用,光合速率、蒸腾速率等指标对水分胁迫响应灵敏,光合作用在植物应对水分胁迫领域的研究极为重要[7-9]。叶绿素荧光技术是以叶绿素荧光特性作为植物光合的有效探针,可以迅速、准确、无损伤地监测环境胁迫状态下植物光合特性,近年来被广泛应用于植物应对水分胁迫响应的研究[10-12]。通常认为,旱涝胁迫会对水稻光合作用产生抑制作用:ASHRAF等[13]认为,干旱、淹涝胁迫可以通过降低光合色素含量、光系统活性、CO2传输水平而抑制光合能力;还有学者发现,干旱条件下水稻叶片净光合速率显著降低,且干旱胁迫越严重,对水稻光合作用的抑制作用越大[14-15]。也有研究认为,适度进行水分胁迫在一定条件下可以促进水稻光合作用,尤其在恢复正常水分条件时可能产生补偿效应:杨伟利等[16]研究发现,短时轻涝下水稻叶片出现光合补偿效应;甄博等[17]研究发现,分蘖期轻度旱涝交替胁迫可以提高水稻叶片中叶绿素含量,增强植株光合能力;郝树荣等[18]认为,水稻生育前期轻度干旱胁迫后复水可以使水稻后期保持较大光合面积。俞双恩等[19]认为,农田水位可以作为水稻控制灌排的调控指标,但以农田水位作为旱涝交替胁迫指标对水稻不同生育阶段叶绿素荧光动力学特性与光合指标的影响研究相对较少。
本文以农田水位作为水稻旱涝胁迫调控技术指标,采用蒸渗测坑控制试验,研究控制灌排条件下旱涝交替胁迫对水稻荧光动力学参数和光合特性的影响,揭示控制灌排条件下旱涝交替胁迫下的水稻生理响应规律,为研究控制灌排条件下作物生理特性和水分胁迫响应机制提供理论依据。
1 材料与方法
1.1 试验区概况
试验于2016年6—10月在河海大学南方地区高效灌排与农业水土环境教育部重点实验室江宁校区节水园区试验场(31°54′N,118°46′E)进行。试验场处于亚热带湿润性气候区,平均气温15.7℃,平均降雨量1 021.3 mm,平均蒸发量900 mm,年无霜期224 d,日照时数2 017.2 h。试验在固定式蒸渗仪测坑内进行,测坑规格为2.5 m×2.0 m×2.0 m(长×宽×深),呈南北向2排布置,每排16个,共计32个,2排测坑中间设地下廊道,地面设2节轨道式电动移动挡雨棚,可根据试验要求进行实时打开或关闭,以准确调控农田水位。测坑内填土为黏壤土,0~30 cm土层土壤容重为1.46 g/cm3,孔隙度为44.97%,田间持水率为25.3%(质量含水率),pH值为6.97,有机质质量比为21.9 g/kg,全氮质量比为0.91 g/kg,速效氮质量比为27.7 mg/kg,全磷质量比为0.32 g/kg,速效磷质量比为12.5 mg/kg。
1.2 试验设计
供试水稻为南粳9108,5月20日泡种,5月22日催芽,5月24日播种,6月20日对蒸渗仪测坑翻耕、泡田、施肥、整平,6月24日移栽,栽种密度为153穴/测坑(30.6穴/m2),每穴3株籽苗。
试验采用双因素试验,即旱涝交替胁迫的阶段:分蘖期、拔节期、抽穗开花期、乳熟期;旱涝交替胁迫的模式:先旱后涝、先涝后旱。共设置8种处理,如表1所示。先旱后涝(HZL)处理,是在生育阶段开始(分蘖期在分蘖开始后7 d)自然耗干田面水层,以田面无水层当日作为受旱起始日,自然耗水至旱胁迫下限后,立即灌水至涝胁迫上限,之后让其自然消退至控制灌排的灌水下限,再按对照(CK)处理水位调控方案进行控制灌排;先涝后旱(LZH)处理,是在生育阶段开始(分蘖期在分蘖开始后7 d)即灌水至涝胁迫上限,让其自然消退,受涝胁迫5 d后排干田面水层,自然耗水至旱胁迫下限,然后灌水至控制灌排的灌水上限,之后按照CK处理水位调控方案进行灌排。所有处理,田面有水层时通过测坑底部排水孔保持2 mm/d的渗漏量,田面无水层时不进行底部排水。除水位调控外,本试验各处理其他农艺措施相同。

表1 测坑水位调控试验处理设计Tab.1 Design of water level control tests on pits mm
注:农田水位以田面为“0”,正值表示田面水层深度,负值表示地下水埋深;“CK”表示对照处理,负值表示灌水下限,正值表示灌水上限;“HZL”表示旱转涝处理,“LZH”表示涝转旱处理,“HZL-1”表示在分蘖期进行先旱后涝胁迫,以此类推;在旱涝交替胁迫阶段,负值表示旱胁迫下限,正值表示涝胁迫上限。
1.3 快速荧光响应曲线模型
叶绿素a快速荧光响应曲线与光合响应曲线具有较高的一致性[20]。应用非直角双曲线模型[21]模拟控制灌排条件下旱涝交替水稻叶片叶绿素快速荧光响应特征,可以得到不同水位调控条件下典型生育阶段水稻叶片潜在最大电子传输速率Emax(μmol/(m2·s))、表观量子效率α(μmol/μmol)、快速荧光响应曲线曲率θ(无量纲)等特征指标。并对曲线初始部分进行线性回归,回归直线与潜在最大电子传输速率水平线的交点为光饱和点(LSP,μmol/(m2·s))。电子传输速率(ETR)计算式为
式中LPAR——光合有效辐射通量密度,μmol/(m2·s)
1.4 测试指标与方法
每个处理分别随机选取3穴非测坑边缘的水稻主茎进行挂牌标号,分别在HZL处理涝胁迫5 d后、LZH处理旱胁迫结束后选择无云或少云的晴天09:30—10:30,测定叶绿素、光合等指标,同时同步测定CK处理的相关指标。测定叶位为:分蘖期、拔节孕穗期水稻最新展开叶中部,抽穗开花期、乳熟期水稻穗叶中部。
使用SPAD-502型叶绿素计测定水稻叶片的相对叶绿素含量(SPAD),选择待测水稻叶片1/2处及其上下3 cm的位置进行测定,以测定的叶位SPAD的平均值作为该处理水稻叶片的SPAD值。
使用MINI-PAM-Ⅱ型超便携式调制叶绿素荧光仪测定水稻叶片光能转化效率Fv/Fm、光化学淬灭系数qP、非光化学淬灭系数qN等指标和快速荧光曲线。测量时将叶夹夹在叶片中部,测定前叶片暗适应15 min,再将光纤传感器插入叶夹,抽开叶夹上的遮光片,开启叶绿素荧光诱导曲线和快速荧光响应曲线功能。测定快速荧光响应曲线时利用仪器自带光量子探头和热电偶记录光合有效辐射通量密度(PAR),分别设定1 500、1 150、850、650、450、300、200、150、100、70、50、0 μmol/(m2·s)共计12个梯度下水稻叶片电子传输速率(ETR),每个光强梯度下测定持续时间为10 s。
使用LI-6400型便携式光合仪测定水稻净光合速率Pn、蒸腾速率Tr、气孔导度Gs和潜在水分利用效率WUEq等指标。
1.5 数据处理与分析方法
采用Excel 2016进行数据统计,OriginPro 2019进行图形绘制和曲线拟合,SPSS 25.0进行数据处理,依据LSD方法进行显著性分析(P<0.05)。
2 结果与分析
2.1 旱涝交替胁迫对水稻叶片SPAD的影响
叶片相对叶绿素含量与其叶绿素含量之间具有明显正相关关系[22],水稻不同生育阶段旱涝交替胁迫对叶片SPAD的影响见表2。由表可知,2种旱涝交替胁迫模式均降低了叶片SPAD,先旱后涝胁迫的降幅更大,HZL-1、HZL-2、HZL-3处理较CK分别显著(P<0.05)降低8.3%、7.3%、3.9%,这种抑制效果随生育期的推进逐渐减小;相应地,先涝后旱胁迫水稻SPAD整体上低于CK,在拔节孕穗期达到显著(P<0.05)水平。

表2 旱涝交替胁迫下水稻叶片的SPADTab.2 Effects of alternate waterlogging and drought stresses on SPAD of rice leaves
注:同列不同字母表示同一生育阶段不同处理间差异显著(P<0.05),下同。
2.2 旱涝交替胁迫对水稻荧光动力学参数的影响
2.2.1水稻叶片Fv/Fm对旱涝交替胁迫的响应
Fv/Fm是叶绿素PSⅡ反应中心最大光化学量子产量,反映PSII反应中心内的光能转化效率[11]。由图1(图中不同小写字母表示同一生育阶段不同处理间差异显著(P<0.05),下同)可以看出,各处理水稻叶片Fv/Fm在不同生育阶段均表现出倒“V”形变化趋势,在拔节孕穗期达到峰值。分蘖期2种旱涝交替胁迫均增加了水稻叶片Fv/Fm,其中LZH-1处理显著(P<0.05)高出CK 4.95%。相应地,其他生育阶段先旱后涝胁迫与CK相比,HZL-2、HZL-3、HZL-4处理叶片Fv/Fm显著(P<0.05)降低了4.56%、5.98%、4.83%,先旱后涝胁迫只有LZH-3处理较CK差异显著(P<0.05),产生了3.32%的降幅。对比同一生育阶段不同旱涝交替模式可以发现,先涝后旱胁迫较先旱后涝胁迫水稻叶片Fv/Fm高出2.01%~4.22%。

图1 旱涝交替胁迫下水稻叶片Fv/Fm的变化Fig.1 Changes of Fv/Fm of rice leaves under alternate waterlogging and drought stresses
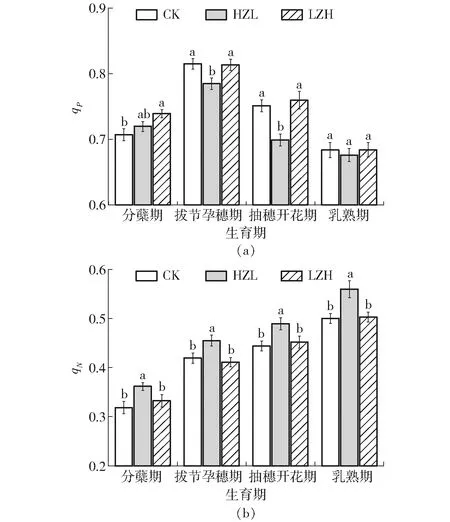
图2 旱涝交替胁迫下水稻叶片qP、qN的变化Fig.2 Changes of qP and qN of rice leaves under alternate waterlogging and drought stresses
2.2.2水稻叶片qP、qN对旱涝交替胁迫的响应
光化学淬灭系数qP用于描述PSⅡ天线色素吸收的光能用于光化学电子传递的份额,反映PSⅡ反应中心的开放程度[11]。由图2a可以看出,qP整体表现趋势与Fv/Fm相似,随水稻生长发育也呈现出倒“V”形趋势,于拔节孕穗期达到峰值。HZL-1处理qP较CK提高1.79%,HZL-2、HZL-3处理分别显著(P<0.05)降低3.72%、6.92%,HZL-4处理较CK无显著差别;LZH-1处理qP较CK显著(P<0.05)提高4.53%,LZH-2、LZH-3、LZH-4处理对qP影响不大。对比处理间差异,先涝后旱胁迫模式较先旱后涝胁迫模式水稻叶片qP提高了1.18%~8.68%。
非光化学淬灭系数qN反映植物将多余光能以热量形式消散的能力,是光合系统的自我保护机制[11]。分析图2b可知,随着水稻的生长发育,水稻叶片qN逐渐提高。先旱后涝胁迫在全生育期对水稻叶片qN影响显著,HZL-1、HZL-2、HZL-3、HZL-4处理qN较CK显著(P<0.05)产生8.50%~12.60%的增幅,而先涝后旱胁迫对其影响不显著。
2.3 旱涝交替胁迫对水稻叶片快速荧光响应特征的影响
2.3.1水稻叶片快速荧光响应曲线对旱涝交替胁迫的响应
不同生育阶段旱涝交替胁迫下水稻叶片快速荧光响应曲线见图3,弱光下叶片PSⅡ反应中心对光照强度的变化响应敏感,ETR随PAR的增加逐渐增大。当PAR处于400~800 μmol/(m2·s)之间时,ETR增速逐渐变小。PAR大于800 μmol/(m2·s)后,ETR趋于平稳。由图3可知,分蘖期(图3a)和拔节孕穗期(图3b)在PAR大于400 μmol/(m2·s)时,旱涝交替胁迫下叶片快速荧光响应曲线均优于CK;相对而言,抽穗开花期(图3c)和乳熟期(图3d)旱涝交替胁迫叶片快速荧光响应曲线低于CK。分析2种旱涝交替胁迫模式发现,相同光照强度条件下,先旱后涝胁迫水稻叶片电子传输速率明显高于先涝后旱胁迫。
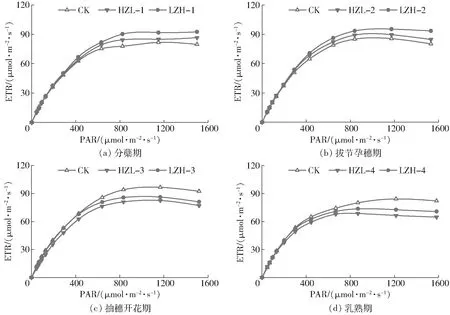
图3 旱涝交替胁迫下水稻叶片快速荧光响应曲线Fig.3 Rice leaf rapid light-response curves of fluorescence under alternate waterlogging and drought stresses
2.3.2快速荧光响应曲线特征参数对旱涝交替胁迫的响应
应用非直角双曲线模型对快速荧光响应曲线(图3)进行拟合,特征参数拟合结果如表3所示,决定系数R2均在0.99以上,均方根误差(RMSE)在1.03~2.67 μmol/(m2·s)之间,说明非直角双曲线模型可以较好地描述旱涝交替胁迫水稻叶片快速荧光响应曲线。潜在最大电子传输速率、表观量子效率、光饱和点等模型特征参数均具有明确的生物意义,比较不同处理的拟合参数可以更加全面地揭示水稻叶片荧光响应特性[23]。
由表3可知,分蘖期旱涝交替胁迫提高了叶片潜在最大电子传输速率,LZH-1、HZL-1处理Emax

表3 旱涝交替胁迫水稻叶片快速荧光响应曲线特征参数Tab.3 Characteristic parameters of rice leaf rapid light-response curve of fluorescence under alternate waterlogging and drought stresses
分别较CK高出15.10%、6.35%;光饱和点LSP由大到小依次为LZH-1、CK、HZL-1,先涝后旱胁迫增强了叶片在强光下的适应能力,先旱后涝胁迫抑制了叶片对强光的适应能力。拔节孕穗期LZH-2、HZL-2处理Emax分别较CK高出13.21%、4.69%,LSP分别高出10.37%、1.57%,表明拔节孕穗期旱涝交替胁迫可以有效改善叶片荧光响应特征,先涝后旱胁迫的改善效果更加明显。
抽穗开花期和乳熟期2种旱涝交替胁迫模式水稻叶片Emax均大幅度低于CK,旱涝交替胁迫降低了叶片的光合能力,先旱后涝胁迫较先涝后旱胁迫产生的抑制作用更大。抽穗开花期和乳熟期旱涝交替胁迫都降低了LSP,抑制了叶片对强光的利用,但处理间对比发现,与CK相比,HZL-3处理LSP降幅较LZH-3增大7.15个百分点,而HZL-4处理较LZH-4降低3.71个百分点,其差异可能由于水稻不同生育阶段需水特性的不同而产生。
2.4 旱涝交替胁迫对水稻光合特性的影响
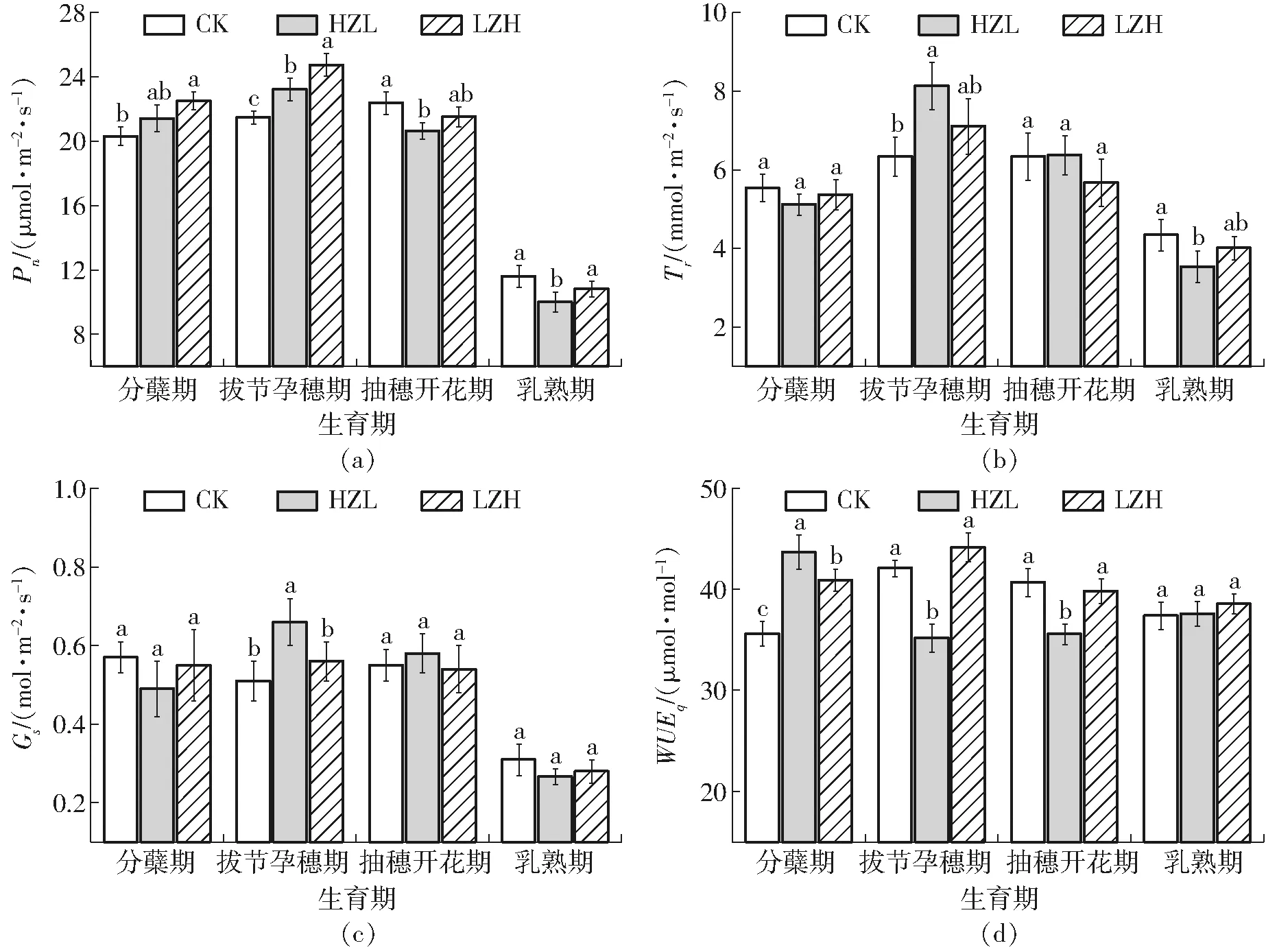
图4 旱涝交替胁迫水稻叶片光合与蒸腾指标的变化Fig.4 Changes of photosynthesis and transpiration index of rice leaves under alternate waterlogging and drought stresses
不同生育阶段旱涝交替胁迫下水稻叶片光合与蒸腾指标的变化见图4。与CK相比,分蘖期旱涝交替胁迫水稻Pn会产生不同程度的增幅,HZL-1、LZH-1处理分别较CK增加5.4%、10.8%,说明分蘖期旱涝交替胁迫对水稻光合作用有一定促进作用。由图4b和图4c可知,Tr和Gs的变化相似,分蘖期旱涝交替胁迫降低了Tr和Gs,可能抑制了水稻蒸腾作用。HZL-1、LZH-1处理WUEq分别较CK显著(P<0.05)增加22.6%、14.9%,促进了叶片潜在水分利用效率。拔节孕穗期HZL-2处理Pn、Tr、Gs较CK分别显著(P<0.05)增加8.2%、28.4%、29.4%,WUEq显著(P<0.05)降低16.4%;LZH-2处理Pn显著(P<0.05)增加15.2%,Tr、Gs、WUEq分别增加12.1%、9.8%、4.9%。表明拔节孕穗期旱涝交替胁迫均对水稻叶片净光合速率、蒸腾速率和气孔导度起到了一定促进作用,先旱后涝胁迫抑制了叶片潜在水分利用效率,先涝后旱胁迫对其影响不大。抽穗开花期HZL-3处理Pn、WUEq较CK分别显著(P<0.05)降低7.8%、15.9%,Tr、Gs与CK差异不大;LZH-3处理Pn、Tr、Gs、WUEq均低于CK,差异均不显著。表明水稻在抽穗开花期经旱涝交替胁迫后叶片光合作用受到抑制,而先旱后涝胁迫对叶片净光合速率和潜在水分利用效率产生的抑制作用更强。乳熟期HZL-4处理Pn较CK显著(P<0.05)降低13.8%,Tr显著(P<0.05)降低8.8%,Gs降低13.8%,LZH-4处理Pn、Tr、Gs均低于CK,差异均不显著,且两种旱涝交替胁迫WUEq较CK处理无显著差异,表明乳熟期旱涝交替胁迫对水稻叶片净光合速率、蒸腾速率、气孔导度表现出了抑制作用,且先旱后涝胁迫较先涝后旱胁迫抑制作用更强,2种旱涝交替胁迫模式对潜在水分利用效率影响不大。
3 讨论
3.1 旱涝交替胁迫对水稻叶片叶绿素含量及荧光参数的影响
叶绿素作为光合作用的重要色素,参与光能的吸收和传递过程。郝树荣等[18]认为水稻分蘖期受旱后复水会明显降低水稻生育后期功能叶片叶绿素的降解,本试验结果表明旱涝交替胁迫降低了水稻叶绿素含量,先旱后涝胁迫由于农田水位从旱胁迫下限直接转入涝胁迫上限,旱、涝胁迫转换过程很快,相对先涝后旱胁迫而言先旱后涝胁迫增加了叶片叶绿素降解,在一定程度上加快了叶片衰老。同时本试验还发现,随着水稻生长发育先旱后涝胁迫对叶片叶绿素含量的抑制作用逐渐变小,表明叶片叶绿素含量对水稻生育前期旱涝交替胁迫较生育后期更为敏感。
分蘖期旱涝交替胁迫对Fv/Fm、qP起到一定补偿作用,其他生育阶段先旱后涝胁迫降低了叶片Fv/Fm、qP,陆红飞等[24]研究发现,分蘖期先旱后涝胁迫并没有降低水稻荧光能力,而拔节期旱涝急转后阻碍了叶片电子传递和能量转移,抑制了光化学活性,与本试验结果具有相似之处。抽穗开花期是水稻营养生长和生殖生长并进的需水关键时期,2种旱涝交替胁迫模式下受涝、旱敏感指数较高[25],本试验发现抽穗开花期旱涝交替胁迫对叶片荧光能力的抑制作用最为明显。对比不同旱涝交替模式结果表明,先旱后涝胁迫水稻叶片Fv/Fm、qP均明显较先旱后涝胁迫降低,差异原因可能先旱后涝胁迫模式存在旱涝急转的过程,水稻从旱胁迫直接淹水转入涝胁迫,相对于先涝后旱胁迫模式涝胁迫缓慢进入旱胁迫来说,对水稻根系活性、叶绿素含量、光合相关酶活性等因素产生更为不利的影响。水稻不同生育阶段先旱后涝胁迫叶片qN高于其他处理,表明叶片PSⅡ反应中心储存的光能更多地以热量的形式消散,从而降低了叶片光合效率,不利于水稻有机物质的积累和产量形成。
3.2 旱涝交替胁迫对水稻叶片快速荧光响应曲线及特征参数的影响
2种旱涝交替胁迫模式可以显著影响水稻不同生育阶段叶片荧光响应特征。随着光强的增加,电子传输速率逐渐增加,当光强达到饱和点后,ETR趋于平稳,这与前人研究结果相似[26-27]。本试验研究发现,光强饱和后,叶片快速荧光响应曲线(图3)略有下降,可能原因是测定快速荧光曲线时光强诱导时间较短,叶片无法快速适应高光强而降低PSⅡ系统的电子传递活性,出现光抑制的现象。
应用非直角双曲线模型对叶片快速荧光响应过程进行模拟,结果显示分蘖期、拔节孕穗期旱涝交替胁迫均对Emax起到一定的补偿效应,且先涝后旱胁迫的补偿作用更强,抽穗开花期和乳熟期旱涝交替胁迫均降低了Emax和LSP,说明水稻生育前期可以适应本试验旱涝交替的环境,而生殖生长开始后旱涝交替胁迫降低叶片光合潜力,不利于叶片对强光的利用。抽穗开花期是有机物质向穗部运输的关键时期,先旱后涝胁迫严重妨碍了根系吸收、矿物营养运输过程,导致叶片对光强变化响应迟钝,表观量子效率降低,所以抽穗开花期先旱后涝后涝胁迫LSP高于先涝后旱胁迫。
3.3 旱涝交替胁迫对水稻光合特性的影响
陆红飞等[27]通过水稻盆栽试验研究发现,分蘖期和拔节期进行水分胁迫,水稻光合表现出明显的补偿效应,甄博等[28]从微观角度研究发现分蘖期旱涝交替胁迫可以促进水稻根系通气组织的形成,提高根系输水能力。本试验研究发现分蘖期和拔节孕穗期旱涝交替胁迫可以促进叶片光合,而抽穗开花期和乳熟期旱涝交替胁迫降低了叶片光合作用水平。同时本研究还发现,不同生育阶段水稻先旱后涝胁迫模式的Pn均低于先涝后旱胁迫,在光合速率的角度进一步说明了先涝后旱胁迫模式优于先旱后涝。分蘖期、乳熟期旱涝交替胁迫在一定程度上抑制了水稻蒸腾作用,拔节孕穗期水稻新陈代谢旺盛,此生育阶段气温较高,旱胁迫后淹水状态下稻田供水充足,水稻通过增大气孔导度,提高叶片蒸腾速率以降低叶温[29],所以先旱后涝胁迫水稻的蒸腾作用得到了增强。分蘖期旱涝交替胁迫可以显著提高叶片潜在水分利用效率,拔节孕穗期、抽穗开花期先旱后涝胁迫降低了叶片潜在水分利用效率,与陆红飞等[27]研究结果略有差异,可能与水分胁迫方式的差异有关。
由于水稻有机干物质形成与累积、向穗部的运输与分配过程主要发生在生育后期[30-32],抽穗开花期、乳熟期旱涝交替胁迫对水稻多数光合生理指标产生逆向影响,严重妨碍了水稻生理生长,先旱后涝胁迫尤为明显,因此在实际稻田水分管理过程中应尽量避免水稻生育后期重度的旱涝交替胁迫,尤其要避免旱涝急转。
4 结论
(1)在水稻不同生育阶段进行旱涝交替胁迫,结果表明,先旱后涝胁迫水稻相对叶绿素含量(SPAD)、最大光化学量子产量Fv/Fm、光化学淬灭系数qP低于先涝后旱胁迫。不同生育阶段旱涝交替胁迫降低了叶片SPAD,分蘖期旱涝交替胁迫水稻Fv/Fm、qP超过对照水平,而在拔节孕穗期、抽穗开花期、乳熟期先旱后涝胁迫呈现出相反的结果。先旱后涝胁迫提高了水稻非光化学淬灭系数qN,从而降低了光合效率。
(2)旱涝交替胁迫可以改变水稻叶片荧光响应特征,分蘖期和拔节孕穗期旱涝交替胁迫均对Emax产生一定的补偿效应,抽穗开花期、乳熟期旱涝交替胁迫均降低了Emax和LSP。2种旱涝交替胁迫模式间对比显示,先旱后涝胁迫水稻Emax均低于先涝后旱胁迫。
(3)分蘖期和拔节孕穗期旱涝交替胁迫可以提高Pn,而抽穗开花期、乳熟期旱涝交替胁迫降低了Pn。分蘖期、乳熟期旱涝交替胁迫抑制了水稻蒸腾作用,拔节孕穗期先旱后涝胁迫水稻的蒸腾作用得到了增强。分蘖期旱涝交替胁迫可以显著提高WUEq,拔节孕穗期、抽穗开花期先旱后涝胁迫降低了WUEq。
猜你喜欢
中国农学通报(2022年29期)2022-11-25
水科学进展(2022年4期)2022-09-14
中国农业科学(2022年8期)2022-05-16
灌溉排水学报(2021年2期)2021-03-17
水利规划与设计(2020年1期)2020-05-25
人民长江(2019年12期)2020-01-02
现代农业科技(2018年22期)2018-01-15
吉林农业·下半月(2017年7期)2017-07-15
绿色科技(2017年7期)2017-05-12
农民致富之友(2017年6期)2017-05-10
