
草莓果生刺盘孢菌的生物学特性及致病性测定
2020-01-02 06:49宋丽丽张丽勍高清华
上海农业学报 2019年6期
宋丽丽,张丽勍,高清华,段 可*
(1上海海洋大学食品学院,上海201306;2上海市农业科学院林木果树研究所,上海201403)
草莓炭疽病是严重威胁草莓生产的重要病害之一,发生普遍,分布广泛,主要危害草莓的叶片、叶柄与匍匐茎,严重时病菌侵入短缩茎,致使整株凋萎、枯死[1]。草莓炭疽病属于高温病害,已成为中国南方草莓产区育苗的主要障碍[2-3]。2012年夏季,湖北省草莓炭疽病平均发病率为41.7%,严重田块发病率高达 69%[4]。
胶孢炭疽菌(Colletotrichum gloeosporioides)是引起草莓炭疽病的病原菌之一。胶孢炭疽菌为复合种,Weir等[5]通过多基因联合建树的方法将胶孢炭疽菌复合种划分为22个种和1个亚种。果生刺盘孢菌(C.fructicola)是胶孢炭疽菌复合种之一。Han等[6]研究发现,果生刺盘孢菌极易侵染草莓的叶片和叶柄,接种后30 d,草莓植株的死亡率达到77.8%。目前,国内对果生刺盘孢菌的研究多限于种的鉴定及其遗传结构[7-9],而针对果生刺盘孢菌的生物学特性及其防治的研究鲜有报道。本试验对分离自上海和安徽地区,侵染草莓植株不同部位的果生刺盘孢菌菌株的生物学特性及其致病性进行分析和比较,以期初步了解果生刺盘孢菌的生长习性和致病机理,为进一步研究近年草莓炭疽病流行成因及综合防治提供参考。
1 材料与方法
1.1 材料
供试菌株分离自上海市和安徽省具有炭疽病典型症状的草莓病株,菌株编号及来源信息详见表1。

表1 供试草莓炭疽菌菌株信息Table 1 Information of the tested strawberry anthrax strains
接种草莓材料为来自上海市农业科学院林木果树研究所经过茎尖脱毒的组培苗‘章姬’(易感病)和‘申阳’(抗病)。
1.2 草莓炭疽病病原菌的分离与纯化
将病样采回后消毒处理,组织分离参考陈瑶等[10]的方法,处理后将组织置于加有氯霉素的PDA平板上,28℃暗培养3 d。切取菌落边缘菌丝置于PDA液体培养基中,在摇床中28℃、200 r/min培养1 d。单孢划线分离病原菌参考张昊等[11]的方法。单孢菌转接到PDA平板上于28℃培养用于鉴定。
1.3 草莓炭疽病病原菌的鉴定
1.3.1 菌株形态特征观察
对上海及安徽地区分离到的炭疽病菌进行形态观察及比较。用打孔器沿菌落边缘打取直径为6 mm的新鲜菌饼,将菌饼转接至PDA平板中央,于28℃恒温培养5 d后观察记录菌饼直径,每个菌株培养并观察记录5皿。参考Cox等[12]的方法摇菌配制适当浓度的孢子悬浮液,在显微镜下观察分生孢子及附着孢形态,每菌株至少观察5个视野,重复3次。
1.3.2 分子鉴定
病原菌DNA的提取:刮取新鲜菌丝在液氮中磨碎,收集100 mg粉末,使用OMEGA E.Z.N.A.TMFungal DNA Miniprep Kits提取菌株基因组DNA。基因选择及PCR扩增参考Weir等[5]的方法略有改动,退火温度为ITS(55℃)、CAL(62℃)、ACT(61℃)、GAPDH(61℃)、ApMat(61℃),胶孢炭疽菌复合种种类划分所用引物见表2。取5μL PCR产物,用1.5%的琼脂糖凝胶电泳进行检测。PCR扩增产物序列测定委托生工生物工程(上海)有限公司进行。

表2 试验所用引物Table 2 Primers used in the experiment
将测序结果经序列比对分析和手动修正,按照ITS-CAL-ACT-GAPDH-ApMat的顺序首尾相连。下载GenBank中同时含有这5个基因的炭疽菌属菌株序列及相关模式菌株序列[18](表3),同样按照ITS-CALACT-GAPDH-ApMat的顺序首尾相连后进行分析。参照李河等[8]方法,选取C.hippeastri作为外群,使用MEGA 7.0软件,采用最大似然法(ML)构建多基因序列的系统发育树。
1.4 生物学特性分析
1.4.1 产孢量及孢子萌发测定
参考张敬泽等[19]的方法稍有改动,将培养7 d的菌落用直径为10 mm的打孔器沿边缘打孔,挑取8个菌块置于30 mL PDA液体培养基中,在摇床28℃、200 r/min摇菌3 d。吸取20μL于载玻片上,在显微镜10×镜下观察记录5个视野的产孢量。同上将孢子悬浮液制成1×106个/mL,取30μL于载玻片上,使孢子悬浮液的浓度为在显微镜10×镜视野下每个视野含有45—50个分生孢子。放入垫有湿润滤纸的培养皿中,置于黑暗条件下28℃恒温培养箱中培养,每处理重复3次,在不同时间(3 h、6 h、24 h)观察和统计孢子萌发率(孢子萌发率=已萌发的孢子数/总孢子数)。
1.4.2 温度对各菌株菌丝生长的影响
用无菌打孔器沿菌落边缘打取直径为6 mm的新鲜菌饼,将菌饼转接至新的PDA平板中央后分别置于8个不同温度梯度(5℃、10℃、15℃、20℃、25℃、30℃、35℃、40℃)的恒温培养箱中培养。5 d后采用十字交叉法测量菌落直径。每个处理3皿,重复3次。
1.4.3 pH对各菌株菌丝生长的影响
准备 1mol/LNaOH和1mol/LHCL溶液,将 PDA培养基的 pH分别调至3、4、5、6、7、8、9、10共8个梯度,用无菌打孔器沿菌落边缘打取直径为6 mm的新鲜菌饼,将新鲜菌饼置于不同pH的PDA平板中央,于28℃恒温培养,5 d后采用十字交叉法测量菌落直径。每个处理3皿,重复3次。
1.4.4 不同碳、氮源对各菌株菌丝生长的影响
参考陈秀蓉等[20]的方法,将直径为6 mm的新鲜菌饼置于加有20 g/kg不同碳源的PDA培养基中央,或加有2.5 g/kg不同氮源的PDA培养基中央,以不加任何碳源或氮源为对照,于28℃恒温培养,5 d后采用十字交叉法测量菌落直径。每个处理3皿,重复3次。
1.5 致病性测定
选取经过茎尖脱毒的组培草莓苗‘章姬’(易感病)和‘申阳’(抗病),生根炼苗驯化后,移栽于基质中,在温室培养1个月后,转至HZLED-1000C型智能人工气候箱(上海皓庄仪器有限公司),适应1周后采用喷雾法接种。将不同菌株分离获得的分生孢子制成1×106个/mL的孢子悬浮液,均匀喷洒于草莓叶片上(以有水滴流下为度),以喷洒无菌水为对照[21-23]。每处理5株,3次重复。接种后的草莓植株置于培养箱(25℃)中黑暗覆膜保湿培养24 h,之后揭膜继续正常培养(25℃,L/D=12 h/12 h)。病情分级与不同菌株致病性等级的划分参考王丰等[24]。
1.6 数据分析
利用SPSS 16.0和Excel 2010软件进行生物统计分析,差异显著性检测采用邓肯氏新复极差法进行;使用MEGA 7.0软件构建系统进化树。
2 结果与分析
2.1 形态特征观察
分离自上海及安徽草莓主栽区的6个炭疽菌菌株在PDA上培养5 d后,菌落形态存在一定差异。GQHAH1、GQHAH6、GQH26、GQH133、GQH131菌株菌落隆起,整个菌落疏松、絮状,中心深灰或灰绿色,边缘灰白整齐。GQHAH4菌株菌落较扁平、整个菌落较致密、绒毛状、白色、边缘整齐。分生孢子整体均呈长椭圆形或一端略尖,光滑,无色,分生孢子在水中萌发产生芽管后形成梨形、卵形等暗褐色的附着胞,胞内分布黑色颗粒状物,附着胞形态较稳定,大小不均一(图1)。
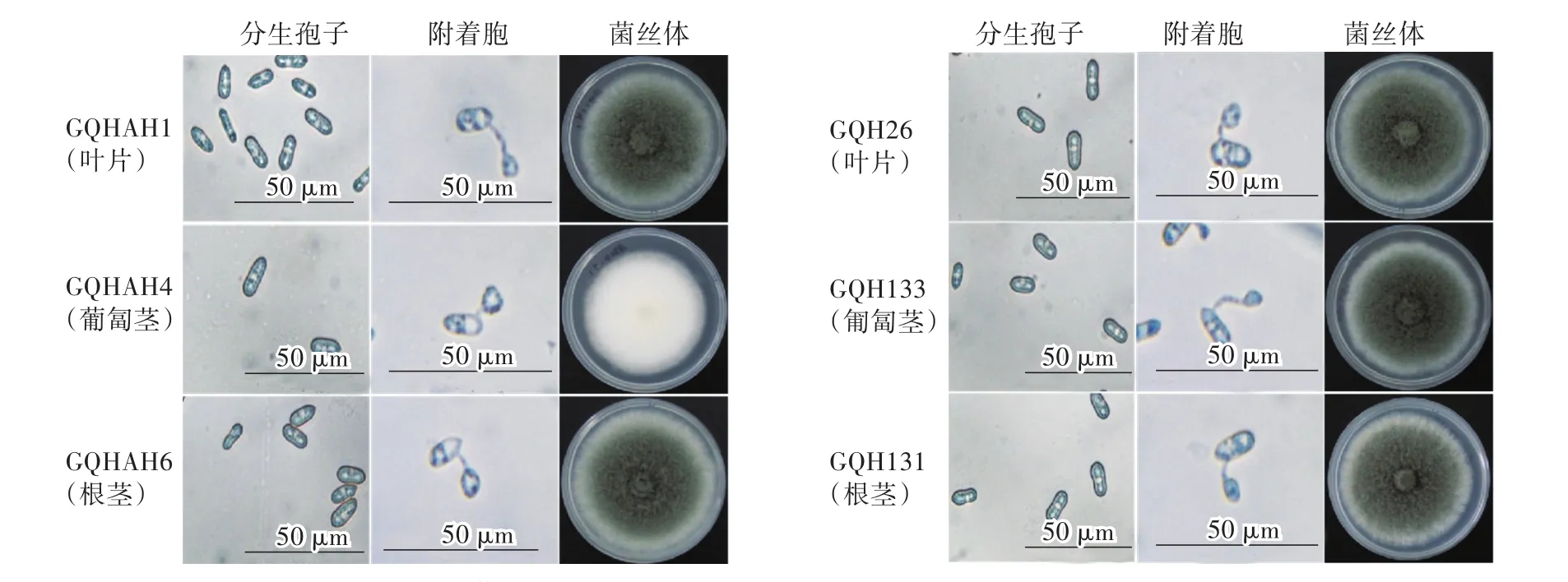
图1 各菌株在PDA培养基(pH=6.8)上生长5 d的菌落形态及40×镜下观察的孢子和附着胞形态Fig.1 Colony morphology of different strains after cultivation on PDA medium(pH=6.8)for 5 daysand themorphology of spore and appressorium observed under 40×m icroscope
根据吴文平等[25]和 LIU等[26]对真菌的分类依据,初步鉴定该病原菌为胶孢炭疽菌(C.gloeosporioides)。
2.2 菌株的分子鉴定
以6个胶孢炭疽菌菌株基因组DNA为模板,扩增ITS(615 bp)、ACT(316 bp)、GAPDH(308 bp)、CAL(756 bp)、ApMat(1 000 bp)基因,得到特异性条带,符合测序要求(图2)。

图2 6个菌株的基因(ITS,ACT,GAPDH,CAL,ApMat)鉴定Fig.2 Identification of genes(ITS,ACT,GAPDH,CAL,ApMat)of 6 strains
使用5个基因(ITS、ACT、GAPDH、CAL、ApMat)的引物,分别扩增供试的6个病原菌,将PCR产物进行测序后,采用多基因联合建树法对其进行比较分析。结果显示,6个草莓炭疽菌菌株与已报道的C.fructicola菌株号为 LC2924、LF131、LC2925、LF132、LC3670、ICMP10642、LF900、LC3368、LF590的菌株处在同一分支上,与已报道鉴定为C.fructicola的模式菌株ICMP18581及CBS130416归在一个种群内(图3)。多基因系统发育树的结果结合病原菌形态特征分析表明,GQHAH1、GQHAH4、GQHAH6、GQH26、GQH133、GQH131病原菌均为草莓果生刺盘孢菌(C.fructicola)。
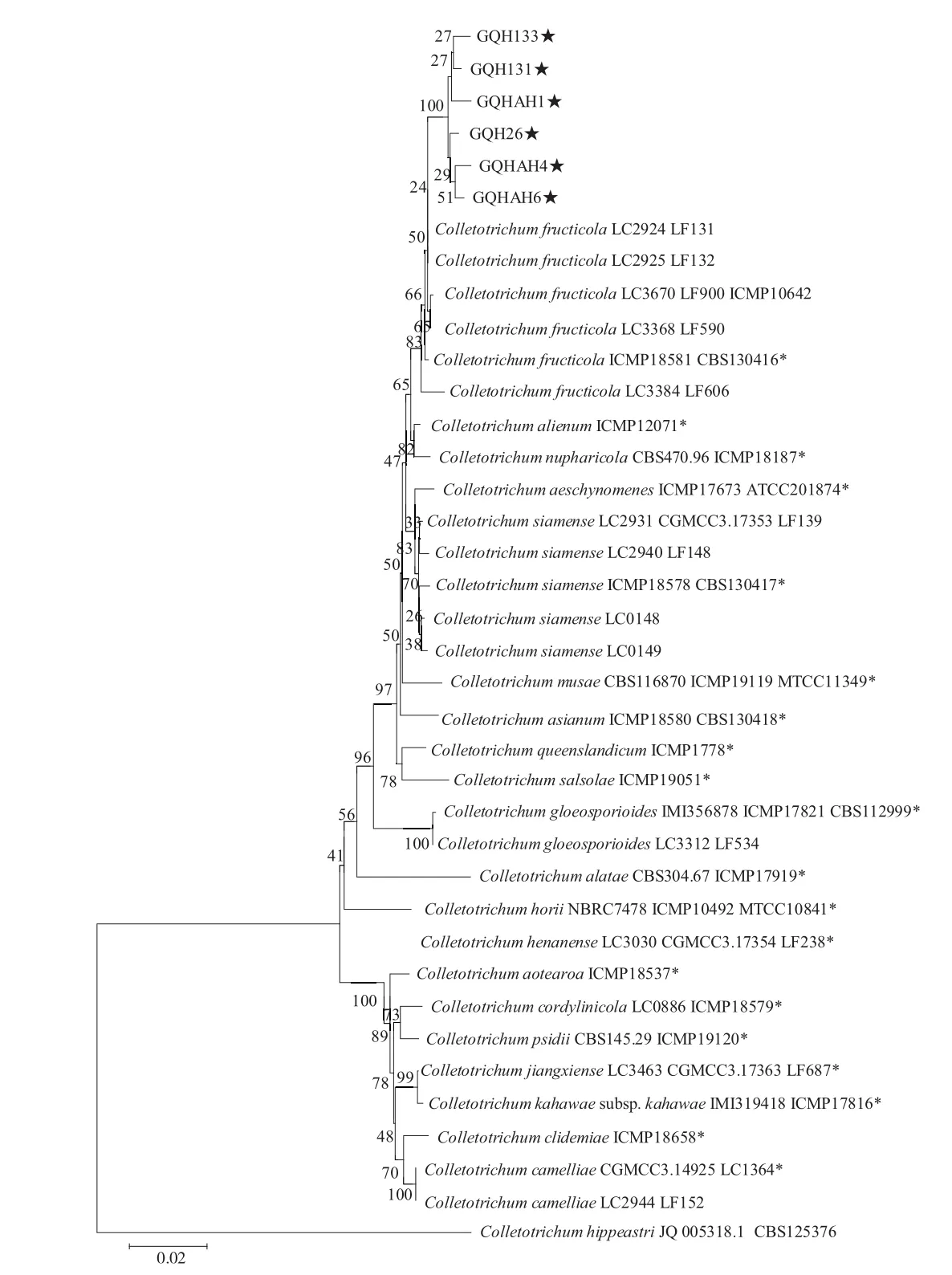
图3 基于ITS、ACT、GAPDH、CAL和ApMat基因联合构建炭疽菌的系统发育树Fig.3 Phylogenetic analysis of anthrax based on ITS,ACT,GAPDH,CAL and ApMat genes
2.3 果生刺盘孢菌的生物学特性分析
2.3.1 产孢量及孢子萌发率比较
如图4所示,各菌株在同等条件下的产孢量存在差异。GQH26菌株摇菌3 d后的产孢量最多,达到291个,GQHAH4产孢量最少,为104个(图4A)。GQH26和GQH131菌株的产孢量显著高于其他菌株。不同菌株在3 h、6 h、24 h的孢子萌发率显著增高,尤其在6—24 h间增长明显。在24 h时,6个果生刺盘孢菌菌株的孢子萌发率接近100%(图4B)。

图4 不同菌株在摇菌3 d后的产孢量(A)及不同时间的孢子萌发率(B)Fig.4 Sporulation capacity of different C.fructicola strains after 3 days’culture(A)and spore germ ination rates(B)at different time
2.3.2 温度对各菌株菌丝生长的影响
如表4所示,在10—35℃范围内,6个果生刺盘孢菌菌株均能生长,当温度低至5℃ 或高至40℃时,菌丝停止生长。当温度为30℃时,不同菌株的菌落直径均达到最大值,表明30℃是较适合草莓果生刺盘孢菌菌丝生长的温度。
当温度为15—35℃时,GQH131与GQHAH1、GQHAH4的菌落直径有显著差异;当温度为10℃ 时,GQH26菌丝生长最快,菌落直径为1.72 cm,GQH133菌丝生长最慢,菌落直径为1.42 cm。由此可见,不同温度对果生刺盘孢菌生长影响不同,这可能与不同菌株对温度的适应不同有关。
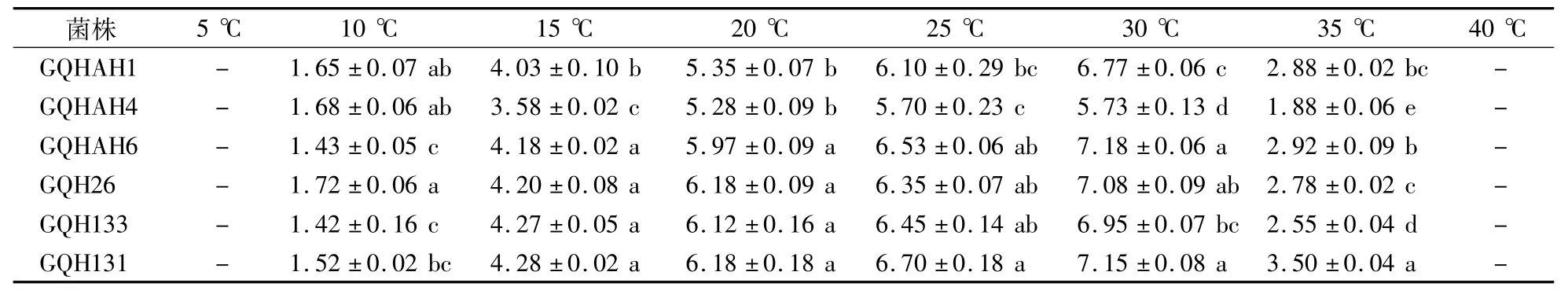
表4 果生刺盘孢菌菌株在不同温度下生长5 d后的菌落直径Table4 Colony diameter of C.fructicola after cultivation for 5 days under different temperature cm
2.3.3 不同pH对各菌株菌丝生长的影响
如表5所示,pH 3—10时,草莓果生炭疽病菌均能生长,而最适菌株生长的pH为6—7。分离自上海的GQH26、GQH133、GQH131菌株在pH为6时菌落直径最大,分离自安徽的GQHAH1、GQHAH4、GQHAH6菌株在pH为7时菌落直径最大。可见,不同pH对不同的草莓果生刺盘孢菌菌株生长影响不同。
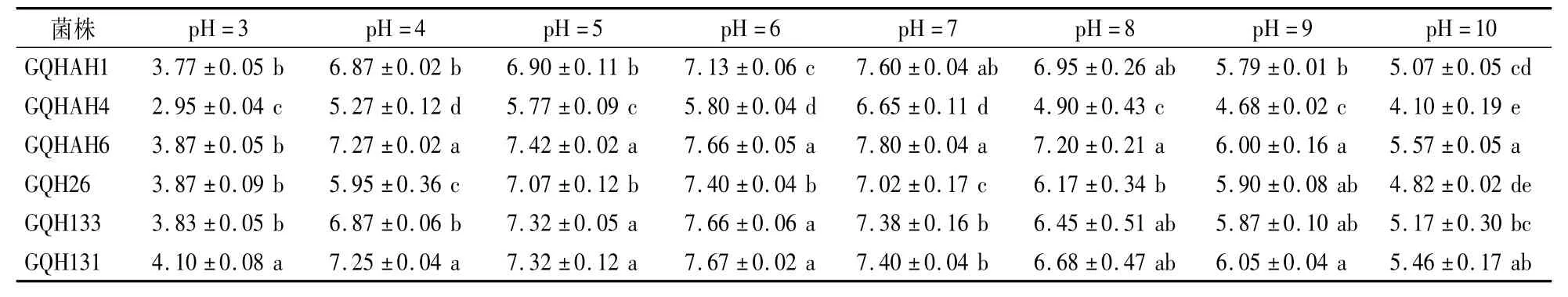
表5 果生炭疽病菌株在不同pH条件下生长5 d后的菌落直径Table 5 Colony diameter of C.fructicola after cultivation for 5 days at different pH cm
2.3.4 碳源对各菌株菌丝生长的影响
供试菌株在葡萄糖、蔗糖、麦芽糖、甘露醇和乳糖为碳源的培养基中均能生长(表6),但菌丝直径存在显著差异。菌株GQH26、GQH131、GQH133、GQHAH1、GQHAH6对碳源的利用偏好顺序为葡萄糖 >蔗糖>麦芽糖>甘露醇>乳糖;GQHAH4对碳源的利用偏好顺序为麦芽糖>蔗糖>葡萄糖>甘露醇>乳糖。同一碳源条件下不同菌株的菌落直径有差异,其中以麦芽糖为碳源时,GQH131菌株菌落直径最大,为7.28 cm,GQHAH1菌株菌落直径最小,为5.92 cm。
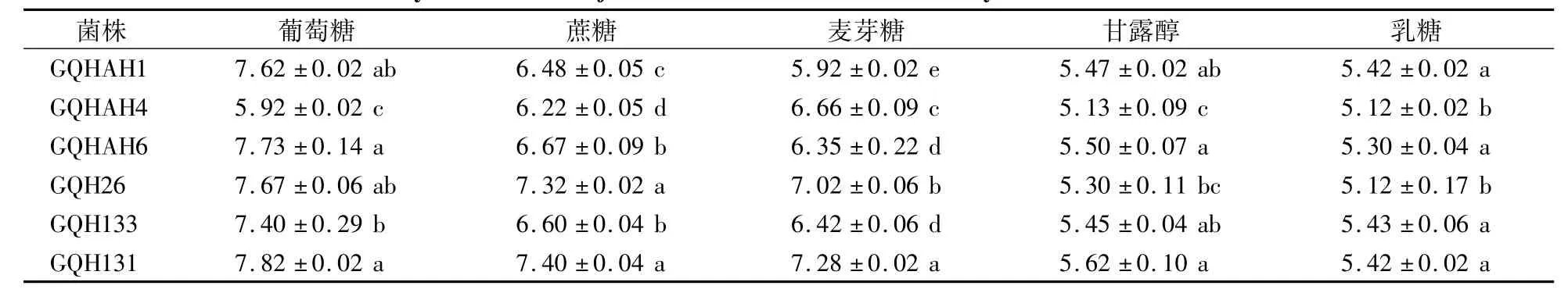
表6 果生刺盘孢菌菌株在不同碳源条件下生长5 d后的菌落直径Table 6 Colony diameter of C.fructicola after cultivation for 5 days w ith different carbon sources cm
2.3.5 氮源对各菌株菌丝生长的影响
如表7所示,菌株GQH26、GQH131、GQH133对氮源的利用偏好顺序为铵盐>丙氨酸>甘氨酸>L-半胱氨酸>尿素;GQHAH1、GQHAH4、GQHAH6对氮源的利用偏好顺序为铵盐>甘氨酸>丙氨酸>L-半胱氨酸>尿素。可见,氮源对不同果生刺盘孢菌菌株菌丝生长的影响存在差异。
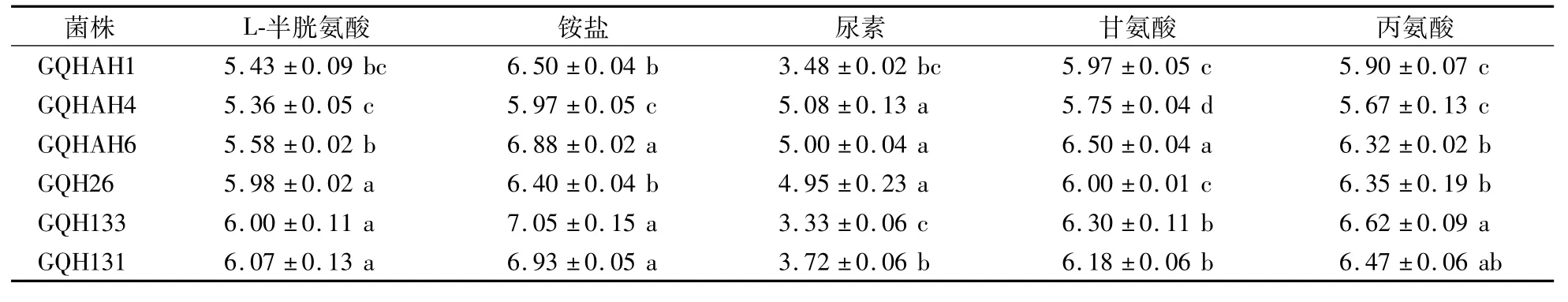
表7 果生刺盘孢菌菌株在不同氮源条件下生长5 d后的菌落直径Table 7 Colony diameter of C.fructicola after cultivation for 5 days w ith different nitrogen sources cm
2.3.6 不同果生刺盘孢菌菌株对草莓叶片的致病性测定
利用不同菌株接种处理易感病草莓品种‘章姬’和抗病品种‘申阳’(苗龄均为30 d),对不同菌株的致病性进行比较。结果表明(图5),GQH131菌株对‘章姬’的致病力显著高于其他菌株,病情指数达到61.98,而其他5个菌株的致病性无显著差异。各菌株对抗病品种‘申阳’的致病性测定表明,GQH131菌株对其致病性显著高于GQHAH1和GQHAH4,病情指数为45.45。可见,来源不同的果生刺盘孢菌菌株致病性存在一定程度的分化。
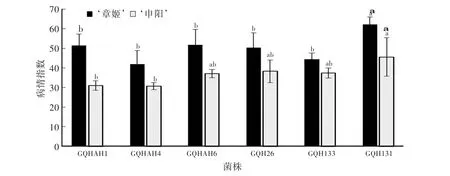
图5 ‘章姬’草莓和‘申阳’草莓接种不同菌株9 d后的病情指数Fig.5 Disease index of‘Akihime’and‘Shenyang’strawberry inoculated w ith different strains for 9 days
3 讨论
草莓是国内外重要的小水果之一,近年来随着全国范围内草莓种植面积的扩大,设施密闭及高温高湿环境等导致草莓炭疽病发生日益严重,造成草莓减产25%—30%[27],严重影响草莓的产量和品质。草莓炭疽病是草莓生产中的一种重要病害,草莓炭疽病的主要病原菌有草莓炭疽菌(C.fragariae)、胶孢炭疽菌(C.gloeosporioides)和尖孢炭疽菌(C.acutatum)[28],均为复合种群,群内种类多,种间菌株变异较大。
传统的真菌分类主要基于形态学特征和ITS序列分析,但对于像炭疽菌属这类近似种之间差异微小,且随着寄主与病原菌之间相互协同进化,形态上的分类标准更为模糊[29]。同时,单一的ITS序列分析对多数近缘种和复合种构建的系统树在一些关键进化节点的支持率较低,不能进行识别[30]。而多基因聚合鉴定比单基因鉴定更加有效,使得基因树更能准确地推测物种,特别是近缘物种的系统关系[31]。采用传统的形态学和ITS序列分析,对于研究胶孢炭疽菌这样的复合种内的遗传关系结果十分不可靠。本试验利用多基因联合建树法对分离自上海和安徽不同组织器官的草莓炭疽病菌进行进一步的分子鉴定,结果显示试验菌株均与果生刺盘孢菌(C.fructicola)归为一个种群。结合病原菌形态特征,鉴定试验所用6个菌株为果生刺盘孢菌。
对果生刺盘孢菌生物学特性分析表明,草莓果生刺盘孢菌对温度、pH等适应范围广,在温度10—35℃及pH 3—10均能生长。较适温度为25—30℃,温度低于5℃或高于40℃时菌丝停止生长。本研究结果与张海英等[32]对草莓炭疽菌生物学特性的研究略有差异,张海英等认为草莓炭疽病病原菌最适温度为25℃,这可能与不同来源及不同菌种的病菌对温度的适应范围不同有关。
本试验发现草莓果生刺盘孢菌最适pH为6—7,与任小杰[33]对草莓炭疽菌的研究、杨成德等[34]对马铃薯炭疽病菌的研究结论一致。此外,本试验表明,果生刺盘孢菌能利用多种碳氮源。在不同碳源中,各菌株对碳源的利用略有差异,其中葡萄糖和蔗糖较适合该菌的生长;在不同氮源中,菌丝在铵盐培养基上生长最快,尿素培养基最慢。
从草莓果生刺盘孢菌接种抗病性不同的草莓品种‘章姬’和‘申阳’叶片后的病情指数来看,不同菌株的致病力存在一定差异。对易感品种‘章姬’的致病性显示,GQH131与其他5个菌株有显著差异;而对抗病品种‘申阳’的致病性仅GQH131与GQHAH1、GQHAH4有显著差异。说明来源不同的菌株在进化过程中,其致病性产生了一定程度的分化,且不同品种的草莓对菌株的抗病性存在差异。
本研究可为生产上防治草莓炭疽病提供参考,当温度为25—30℃ 时,需加强对该病害的防控。Freeman等[35]报道,炭疽病可在无菌的土壤中存活至少1年,这与其广泛的适应性有关。另外,不同草莓果生刺盘孢菌在生物学特性也有一定差异,生产上应该因地制宜地指导不同地区草莓果生刺盘孢菌的科学防控。本研究分离菌株数量有限,关于不同来源的草莓果生刺盘孢菌生物学特性和致病性分化情况,尚待大量收集不同地区的病原菌,结合室内观测和田间小区试验进行深入研究,以此更好地指导田间种植防护工作。
猜你喜欢
安徽农业大学学报(2022年2期)2022-11-09
今日农业(2022年15期)2022-09-20
南京农业大学学报(2022年2期)2022-04-01
山东农业科学(2022年2期)2022-03-12
中国土壤与肥料(2021年5期)2021-12-02
现代畜牧科技(2021年4期)2021-07-21
中国马铃薯(2020年4期)2020-09-10
东北农业大学学报(2020年3期)2020-05-14
中国动物保健(2015年4期)2015-10-21
营销界(2015年23期)2015-02-28
