
湿地松改良种子园无性系的遗传评价及新一轮育种亲本选择*
2020-02-12 06:57邓乐平吴惠姗李晓华廖仿炎李义良郭文冰赵奋成
林业与环境科学 2020年4期
邓乐平 黄 婷 王 哲 吴惠姗 李晓华 廖仿炎 李义良 郭文冰 赵奋成
(1.台山市红岭种子园,广东 台山 529223;2.广东省森林培育与保护利用重点实验室/广东省林业科学研究院,广东 广州 510520)
遗传测定是林木改良工作的中心环节,通过开展遗传测定评价亲本的育种价值、选择新一轮的育种亲本、估计遗传参数和验证或预估遗传增益[1-2]。林木育种中子代测定的交配设计种类多样,每种设计各有利弊,其中,自由授粉交配设计虽然存在父本来源不清、不能估计特殊配合力信息等方面的不足,但该方法具有技术简单易行、成本低等优点,能够提供母本的一般配合力信息,为种子园的去劣疏伐提供依据[1],因而在多个树种的改良活动中受到广泛应用,我国的杉木(Cunninghamia lanceolata)、马尾松(Pinus massoniana)、火炬松(P.taeda)等树种均有关于利用自由授粉子代开展遗传测定的研究报道[3-6]。
湿地松(P.elliottii)原产美国东南部,具有适应性强、早期速生、树干通直、松脂产量高质量好等优点,是重要的建筑材、纸浆材和产脂树种,在世界上多个国家和地区均有引种[7-8]。湿地松于1930 年代引入我国,在我国南部低山丘陵、沿海地区表现优良,目前是我国南方省区主要的商品林树种。我国开展湿地松良种选育始于1964 年[7,9],经过几代人的努力,在广东省台山县(市)建起国内的第一个湿地松初级种子园、一代种子园、改良种子园、精选种子园和二代种子园。其中,湿地松改良种子园生产区建于1988—1997年,面积52 hm2,建园无性系147 个。建园材料来源广泛,包括美国佐治亚州湿地松种子园的优良无性系、台山湿地松初级种子园中经过子代测定后向选择的优良无性系、子代测定林和种源试验林中前向选择的优树无性系。由于多数建园无性系的遗传品质尚未验证,因此,项目组在该种子园中多批次采收无性系的自由授粉种子并开展多地点遗传测定。本文报道早期营建的1 块测定林的测定结果。
1 材料与方法
1.1 材料来源
参试材料为28 个湿地松自由授粉家系。其中,9 个来自台山湿地松一代种子园,19 个来自台山湿地松改良种子园。
1.2 试验地点
试验林营建在台山市红岭种子园试验区内。该园地处22°11′N,112°49′E,属南亚热带海洋性气候,年均温21.8 ℃,年降水量1 940 mm,平均海拔30 m。土壤为花岗岩发育的酸性砖红壤,pH值5.0~5.5,土层80~100 cm。自然条件适合湿地松生长发育、开花结实。该林地前茬为桉树,但土壤肥力较低。
1.3 田间设计
上述家系种子于1999 年12 月育苗。2000 年6 月造林,株行距3 m×3 m。测定林采用随机完全区组设计,8 次重复,5 株单行小区。试验区四周种植2 行保护行。
1.4 生长量调查
分别于造林后第2、3、4、13、14、15、18年对测定林开展年度生长量调查,测量每木的树高(ht,单位为m)和胸径(dbh,单位为cm),并计算单株材积(vol,单位为m3)。材积计算式为:vol=f×π×ht×dbh2/4 。式中,π 为圆周率;f 为形数,取0.5。
考虑到本文的主要目的是评价参试家系的遗传品质和选择新一代的育种材料,将仅利用第4、13、18 年的调查数据作分析。
1.5 数据统计分析
采用SAS 统计分析软件MIXED 过程对生长数据作方差分析[10],估计改良种子园和一代种子园子代的生长性状平均值(lsmean)并作差异显著性比较。单株数据统计分析的线性模型为:yijkl=μ+bi+gj+f(g)k(j)+(bg)ij+(bf(g))ik(j)+eijkl,式 中yijkl为单株观测值,μ为群体平均值,bi为区组效应,gj为种子园效应,f(g)k(j)为种子园内家系效应,(bg)ij为区组与种子园的互作效应,(bf(g))ik(j)为区组与种子园内家系的互作效应,eijkl为误差效应。其中,种子园、种子园内家系为固定效应,其它因素为随机效应。
采用Asreml 统计软件BLUP 方法估计湿地松改良种子园19 个家系的母本育种值和子代个体育种值[11-12]。
采用SAS 统计分析软件CORR 过程的pearson 方法计算母本育种值的年度相关系数[13]。
2 结果与分析
2.1 湿地松子代测定林的总体生长表现
表1 列出测定林3 个测定年龄的生长量。由表1 可见,林分保存率(sur)在4 a 生时为97.40%,到13 a 生时仍然高达92.60%,到18 a生时林分的保存率仅为52.9%。测定林4 a 生时平均树高4.20 m,平均胸径6.76 cm,年平均树高1.05 m、胸径1.67 cm;到13 a 生时,林分平均树高为10.53 m、平均胸径14.91 cm,4~13 a 生间的年均树高、胸径分别为0.70 m、0.91 cm;到18 a生时,树高、胸径年生长量下降更为明显,甚至出现单位面积蓄积量小于13 a 生时的情况。保存率和生长量下降虽然与树种自身的生物学特性有关,但与2014—2018 年间该测定林作生产性的松脂采割有更大的关系。

表1 湿地松子代测定林3 个年龄的生长表现Table 1 The growth performance of progeny test forest of slash pine in three different years
2.2 湿地松改良种子园无性系的现实增益
根据参试家系的来源背景,把来自湿地松一代种子园的9 个自由授粉家系归为一代种子园子代群体,把来自改良种子园的19 个自由授粉家系归为改良种子园子代群体,对3 个年份试验数据作方差分析,结果显示2 个种子园间生长量存在显著甚至极显著的差异(4、13 a 生时,F=20.99~33.80,P<0.000 1;18 a 生时,F=5.15~7.45,P=0.006 8~0.024 2)。改良种子园子代生长量显著大于一代种子园子代(表2),在4、13、18 a 时,改良种子园子代相对于一代种子园的单株材积现实增益分别为25.66%、22.68%和15.08%。说明改良种子园子代生长优势的表现是稳定的,也说明建园无性系选择有效。

表2 湿地松改良种子园子代与一代种子园子代3 个年龄的生长差异比较Table 2 Comparison of growth differences between the progeny of slash pine in the improved seed orchard and the firstgeneration seed orchard in three different years
2.3 湿地松改良种子园19 个无性系的遗传评价
利用BLUP 估计了来自改良种子园的19 个自由授粉参试家系的母本无性系的生长性状育种值(表3),19 个无性系的育种值差异明显,在4 a 生时,树高性状的育种值介于-0.81~1.39 m 之间,13 a 生时介于-2.19~1.70 m 之间,18 a 生时介于-1.95~1.59 m 之间。胸径、单株材积的育种值也显示出大的变异范围。由表中还可见,总体上,母本无性系在3 个年龄的育种值的排序基本一致。4 a 生树高育种值与13 a、18 a 生的育种值相关系数分别为0.835 3、0.740 7,4 a 生单株材积育种值与13、18 a 生时的单株材积育种值的相关系数分别为0.956 9、0.918 5(表4),均达到极显著水平。说明湿地松无性系生长性状的育种值在早、中、后期是相对稳定的。

表3 湿地松改良种子园19 个母本无性系3 个年龄的生长性状育种值Table 3 The breeding values of 19 clones of improved slash pine seed orchard at 3 ages in the improved seed orchard

表4 湿地松改良种子园19 个母本无性系3 个年龄生长性状育种值间的相关系数Table 4 Correlation coefficient between breeding values of growth traits of 19 maternal clones in 3 observation years
改良种子园中,7 个从美国引进的无性系的育种值差异很大,育种值较大的无性系分别为C09、C12、C13,其中无性系C09 的生长表现尤为突出,在参试的19 个无性系中育种值最大,其18 a生时的单株材积育种值为0.186 1 m3,比群体平均值大107.89%。另外4 个无性系的育种值较小,其中无性系C15 的单株材积育种值为-0.092 6 m3,在参试的19 个无性系中最小。说明直接从国外种子园引进无性系有一定的效果,但也存在一定的风险,需要作测定、比较。
来自台山湿地松一代种子园的6 个无性系(后向选择)中,C16 的育种值最大,其18 a 生时的单株材积育种值为0.116 5 m3,比群体平均大67.55%。其他5 个处于中等偏小水平。显示选择群体遗传品质不高或选择的准确性偏低。
早期子代测定林中选择的4 个优树无性系(前向选择)中,C16 的子代C16-4 的单株材积育种值最大,18 a 生时为0.061 9 m3,比群体平均大35.88%;C16-1 的育种值略低于群体平均,C19-2和C25 的育种值更小。比较母本无性系C16、C19与子代无性系C16-1、C16-4、C19-2 的育种值可以发现,育种值大的母本,其子代的育种值也较大,此外,子代的育种值低于母本的育种值。
选自种源试验林的2 个优树无性系,其生长性状育种值在参试的19 个无性系中处于中等或中上水平,其中,无性系C26 的育种值较大,其在18 a 生时的单株材积育种值为0.050 5 m3,比群体平均值大29.28%。
2.4 新一代育种资源选择
利用Asreml 软件估计来自湿地松改良种子园19 个无性系的自由授粉子代的树高、胸径、单株材积育种值,按入选率为3%的比例选择生长表现优良的个体,预期选择优树15 株。在评选优良单株中,本文采用了2 种选择模式,模式一:是根据单株育种值排序,选择单株材积育种值最大的15 株树木;模式二:首先选择生长量大于群体平均值的家系,然后在每个入选家系内选择2~3株育种值最大的个体,共选择优良单株15 株。2种选择模式的评选结果列于表5。由表5 可见,2种选择模式共选择优树17 株,其中有13 株优树在2 种选择模式中同时入选,均来自6 个优良家系。模式一入选的15 株优树中,由于对家系内的单株选择数量没有约束,因此,家系F09 内入选数量达5 株,而家系F16-4、F26 的入选优树仅各有1 株。模式一的入选优树的树高、胸径、单株材积育种值分别介于0.40~1.71 m、6.28~9.71 cm、0.161 6~0.276 9 m3,平均值分别为1.06 m、8.01 cm、0.215 4 m3。模式二的入选优树的树高、胸径、单株材积育种值分别介于0.40~1.71 m、4.53~9.76 cm、0.114 9~0.276 9 m3,平均值分别为1.06 m、7.90 cm、0.209 1 m3。显示这2 种模式的选择效果近似。考虑到育种群体遗传多样性的需要,采用模式二评选优树比较合适。
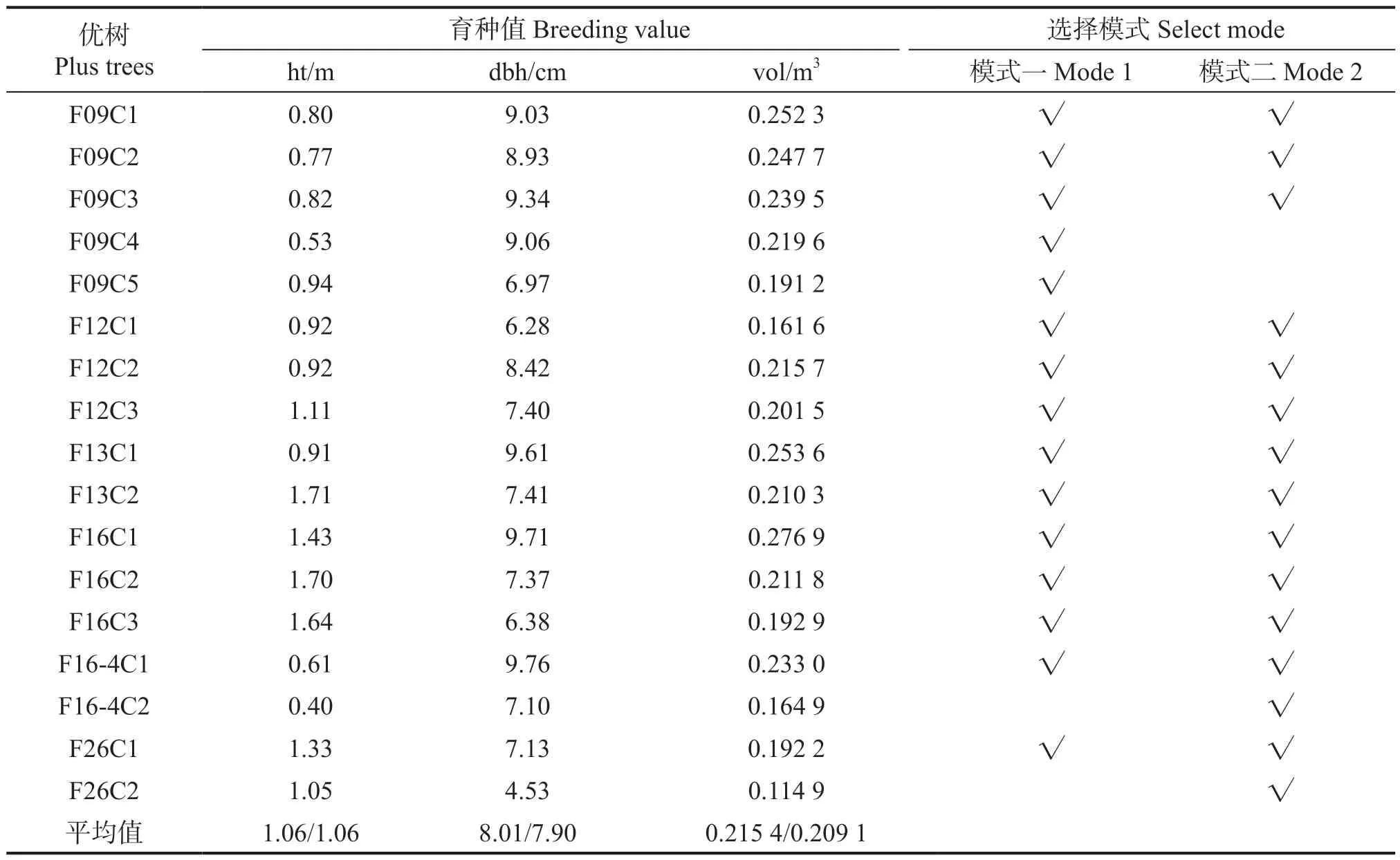
表5 入选湿地松优树18 a 生时的生长性状育种值Table 5 The breeding value of the growth traits of the selected slash pine plus trees at the age of 18
3 讨论与结论
本试验结果显示,与一代种子园的子代比较,改良种子园19 个无性系的子代在3 个测定年龄都表现出明显的生长优势,在4、13、18 a 生时的单株材积现实增益介于15.08%~ 25.66%,该结果与改良种子园的预期材积增益接近[9],由于这19 个无性系的分株数量占了整个湿地松改良种子园的62.79%,其平均表现基本反映了该种子园的总体水平,因此,本结果也表明湿地松改良种子园的遗传品质达到了预期效果。
参试的改良种子园19 个家系的母本无性系中,生长性状育种值较大的无性系有6 个,其中,3 个为直接从美国湿地松种子园引进的无性系C09、C12、C13,1 个为台山湿地松初级种子园的优良无性系C16,1 个为该优良无性系的自由授粉子代C16-4,1 个为1 湿地松种源试验林的优树无性系C26。前5 个无性系遗传品质在其他试验中同样表现优良[14-16],因此,可以继续作为新一轮的育种亲本甚至是核心育种亲本。有多个研究表明,火炬松、湿地松、湿加松等树种的子代测定具有良好的早晚相关,开展早期选择是有效的[17-19]。本研究结果显示,19 个无性系的4、13、18 a 生的生长性状育种值存在极显著的正相关,尤以13 a 生与18 a 生的育种值相关性更为密切,再次证实南方松树开展早期选择是可靠的。
林木的多世代轮回选择育种中,优树选择是重要的育种活动之一,随着林木育种进入高世代,育种群体将主要由来自遗传测定林的优树无性系组成。在高世代育种亲本的选择,要兼顾选择增益与遗传多样性,为了避免出现入选优树仅选自某些优良家系的情况,通常设定入选家系内优良个体的入选比例或数量[20]。基于这方面的考虑,本文设定了整个测定林中优树入选率为3%、每个入选家系内选择2~3 株的限制,根据单株材积育种值从6 个家系中共选择出优良个体15 株。我国现阶段湿地松的重要经济性状主要是材积、松脂产量、木材密度,本文仅针对生长性状评选优良个体,后期还要评价其松脂产量和木材密度,为不同的育种目标提供合适的育种亲本。由于湿地松生长性状与松脂产量或木材密度的遗传相关性不高[21-23],甚至负相关[24],这15 个生长优势突出的个体中,一些个体将可能因其他性状未能达标而被淘汰。
普遍认为,林木存在自交或近交不育和衰退的可能性[1]。因此,在制订林木高世代育种策略中,强调采用双亲谱系清楚的交配设计[25],而不提倡父系来源不清的自由授粉或多系授粉交配设计。现在,随着分子标记技术的发展和完善,育种亲本间遗传关系的鉴别成为了可能[26-28],有人提出从自由授粉子代、多系授粉子代中选择优良个体作为育种亲本的设想[29-30],并付诸实践[28-31]。本研究从自由授粉子代测定林中根据生长性状育种值选择出一批优良个体,下一步要通过分子标记手段,确定入选优树的谱系关系、遗传距离等,为设计新的交配组合,以及种子园无性系排列提供科学依据。
松材线虫病是全球森林生态系统中最具危险性、毁灭性的病害之一,全国因松材线虫病损失的松树累计仍达数十亿株,造成的直接经济损失和生态服务价值损失上千亿元。危害对象包括马尾松、黑松(Pinus thunbergii)、红松(P.koraiensis)、落叶松(Larix olgensis)等松树种类,疫情直接威胁我国近9 亿亩松林资源安全[32]。据报道,广东发生松材线虫病的情况也不容乐观[33-34],但湿地松林分感病率相对较低[34]。同时,研究也发现[35-36],原产于美国的湿地松、火炬松对松材线虫病具有较高的抗性。因此,可以利用我国现有的湿地松、火炬松育种资源,与国内的其他松树开展杂交育种,选育出抗松材线虫病的新品系,提升我国松树人工林的综合效益。
猜你喜欢
浙江林业科技(2022年4期)2022-07-16
农业与技术(2021年6期)2021-12-12
辽宁林业科技(2021年3期)2021-07-08
湖南林业科技(2021年2期)2021-06-04
林业与环境科学(2021年6期)2021-02-11
山西水土保持科技(2020年2期)2020-12-22
农家科技中旬版(2020年7期)2020-08-10
林业与生态(2020年2期)2020-04-16
科学咨询(2020年36期)2020-01-06
福建林业(2016年6期)2016-08-16
