
碳源共施对不同氮源的净化效果的影响
2020-03-09 06:13覃思跃陈国宁陆立海李文文陈永利宋海农
桂林理工大学学报 2020年4期
覃思跃, 陈国宁, 陆立海, 蒋 婷, 李文文, 陈永利, 宋海农
(广西博世科环保科技股份有限公司, 南宁 530007)
氮、 磷含量过高是水体富营养化的主要因素, 目前, 人们已经能够在一定程度上控制水体中的磷含量, 但对水体中氮的去除较为困难, 随着排放量增加, 氮逐渐成为水体富营养化的成因之一[1]。工业上对氮的去除研究较晚,多是通过构筑物中的微生物作用,但传统的生物处理工艺,如膜生物法、A2O等工艺都有处理极限,导致污水厂出水氮浓度偏高,C/N较低[2]。碳源短缺已经成为低C/N比污水中氮去除的限制因子[3-4]。
自然水体中亚硝态氮是铵态氮向硝态氮转化的中间产物, 铵态氮、 硝态氮是无机氮的主要存在形态[5-6]。由于两者的化学物理性质不同, 以及植物对其吸收特性的不同等, 导致两者的去除机理不同[7]。一般认为, 硝化反硝化作用是水体氮的主要去除途径:硝化作用是指铵态氮在好氧条件下发生化学氧化或者好氧铵氧化作用最终转化为硝态氮的过程;反硝化作用是硝态氮在缺氧条件下转化为中间产物亚硝态氮, 最终转化为气体氮化物的过程。但有研究认为, 由于植物根系的泌氧作用, 其周围形成了无数好氧-缺氧的微域, 因此微宇宙在好氧条件下也可以发生硝化反硝化作用, 即好氧硝化反硝化作用[8]; 另外, 植物还可通过向水体中分泌氧气和有机物等作用改变水质, 影响氮的形态转化和去除[9]; 植物根系等为微生物提供附着位点, 促进微生物的生长繁殖, 从而促进硝化反硝化作用的进行[10]。在自然水体中铵态氮和硝态氮经常是同时存在的, 两者之间存在相互转化和相互作用, 影响彼此的净化效果[11]。
植物能够吸收低浓度氮源, 对其具有较好的去除效果。 碳源种类和C/N比均显著影响水体中硝态氮的去除, 但少有研究碳源对铵态氮的去除以及碳源对铵态氮和硝态氮共存去除效果的影响。 本文通过对比是否施加碳源时铵态氮、 硝态氮及铵态氮和硝态氮混合氮源的去除效果, 探究碳源对不同氮源去除效果的影响及其机理, 以期为水体氮净化和防治水体富营养化提供理论依据。
1 材料与方法
1.1 装置和材料
试验装置如图1所示, 以5 L透光玻璃容器为反应器, 用橡胶塞密封, 上端与气泵连接, 并布设采样口和气体采样袋。在反应器底部平铺5 cm厚的土壤, 种植长势一致的矮生苦草, 加入自来水预培养。2周后, 置换为自配污水, 密封好, 置于恒定光照下培养, 光谱为红蓝光4∶1, 光照强度为7 800 Lux。按照设定的采样周期定时采集气体样品、 水样和植物样品待测。其中气体样品用连续通入6 L氩气的方法采集。试验植物矮生苦草, 株高为10~15 cm, 每个反应器20株。

图1 试验装置示意图
1.2 试验方案
主要考虑碳源(无水乙醇)对苦草净化铵态氮、 硝态氮、 铵态氮硝态氮混合氮源效果的影响。自配污水详情见表1。

表1 自配污水碳氮源组成及其浓度
为了排除气体收集过程中O2和CO2变化对植物光合作用和呼吸作用的影响, 每个样品采样一次即结束。其中硝氮和硝氮+碳源处理每6 d取样一次, 其他处理每天取样一次。每个处理均取样11次, 每次取3个平行样品作为重复, 故每个处理设33个重复, 总计198个样品。
1.3 样品取样和分析
1.4 计算指标
系统中产气量主要来自于反硝化作用及厌氧氨氧化作用。产气量大小表明反硝化作用、厌氧氨氧化作用的大小。日产气量等于当日累积产气量减去前一日累积产气量。即日产气量(Qn)为
Qn=Qn,T-Qn-1,T,
式中:Qn为第n天的产气量, mg·L-1·d-1;Qn,T为第n天的累积产气量, mg·L-1;Qn-1,T为第n-1天的累积产气量, mg·L-1。植物氮日净吸收量计算方法与此类似。


2 结 果
2.1 碳源对苦草净化铵态氮效果的影响
当外源氮以铵态氮形态单独施加时, 铵态氮浓度随时间而下降, 去除量为6.18 mg·L-1, 去除率为69.39%(图2a)。总氮去除趋势与铵态氮一致(图 2b), 去除量为3.28 mg·L-1, 去除率为34.49%, 铵态氮净化率ANRR仅为53.11%。从图 2c可知, 硝态氮浓度随时间的延长而逐渐升高, 试验结束时硝态氮浓度由初始的0.16 mg·L-1增至3.73 mg·L-1, 为初始的23.31倍, 铵态氮硝化率ANNR为57.80%。
当外源铵态氮与碳源共施时, 铵态氮去除量为8.27 mg·L-1, 去除率为96.16%, 均较外源铵态氮单独施加时高(图 2a)。总氮的去除量为7.72 mg·L-1(图 2b), 铵态氮净化率ANRR为93.35%。试验第1天, 由于水体的好氧环境, 部分铵态氮发生了硝化作用, 导致硝态氮浓度有少量积累, 较原始浓度增加了0.59 mg·L-1, 随后硝态氮浓度逐渐下降, 铵态氮硝化率为1.71%(图 2c)。COD浓度表现为先快速下降随后上升再下降的波动下降过程(图 2d)。由图2e可知, 外源施加铵态氮使水体的溶解氧浓度迅速由初始的7.97 mg·L-1降至3.67 mg·L-1(第1天), 之后随着铵态氮浓度的下降, 溶解氧浓度逐渐上升。试验第2~6天, 外源铵态氮与碳源共施处理亦使水体DO浓度快速下降, 甚至达到厌氧水平(0.6 mg·L-1), 较相同情况下铵态氮单独施加时降低了3.79~4.70 mg·L-1。试验第7天开始, 外碳源共施处理水体DO浓度逐渐上升并逐渐恢复好氧水平。
相同时期(第2~6天), 铵态氮与碳源共施处理的产气量也较第1天时显著增加(图3a)。系统产气量也逐渐降低。与此不同, 铵态氮单独处理试验第2~6天时的产气量保持在0.1±0.002 mg·L-1·d-1范围内, 变化不大, 且与第1天时差异不显著。试验第7天开始, 外碳源与铵态氮共施处理、 铵态氮单独处理, 系统产气量均较低, 说明反硝化作用、厌氧氨氮化作用变小,可能是由于一方面氨氮浓度下降,另一方面水体溶解氧上升,不利于反硝化作用、厌氧氨氧化作用的条件。
由图3b可知, 外源铵态氮单独施加时, 试验前1 d植物对氮源的净吸收量为5.19~7.47 mg·g-1·d-1, 第3天开始下降, 第6天降至最低值0.17 mg·g-1·d-1, 后期植物净吸收量又开始恢复, 但始终低于初期时的净吸收量。外源铵态氮与碳源共施时, 试验初始前2 d植物对氮源的净吸收量也较高, 且较铵态氮单独施加处理最少增加了1.40倍。随后植物氮净吸收量随时间的延长逐渐下降, 第11天达到最小值1.23 mg·g-1·d-1, 较第1天下降了18.56 mg·g-1·d-1。期间, 植物氮净吸收总量为85.25 mg·g-1, 较铵态氮单独施加时增加了2.94倍。

图2 碳源净化铵态氮对水质指标的影响
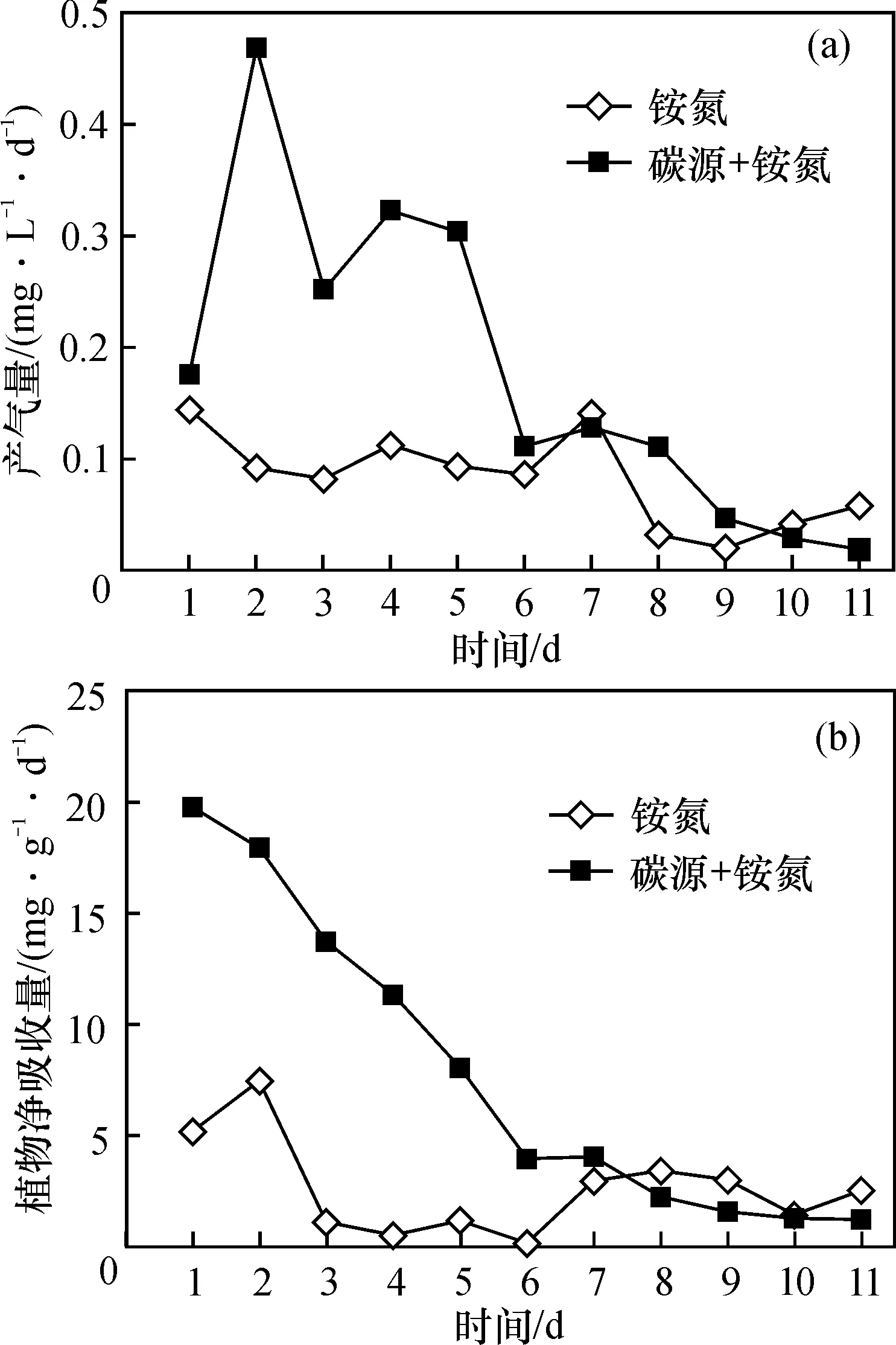
图3 碳源对铵态氮处理系统产气量和植物氮吸收量
2.2 碳源对苦草硝态氮净化效果的影响
由图4a可知, 当外源硝态氮单独施加时, 苦草对硝态氮的去除速率变化过程为: 前6 d慢—第7~12天快—第25~54天慢—第55~66天快。试验期间硝态氮去除总量为1.40 mg·L-1, 去除率为6.83%。总氮的去除变化规律与此相似, 总氮总去除量为1.0 mg·L-1, 去除率为4.79%。当外源碳源硝态氮共施时(图4b), COD浓度表现为先快速下降, 后有所回升, 再下降的波动下降过程。 硝态氮和总氮浓度随时间逐渐下降, 去除量分别为10.17和10.20mg·L-1, 去除率分别为44.71%和44.45%, 均较硝态氮单独处理高。由图4b可知, 试验前12天, 苦草对硝态氮和总氮有很强的去除能力, 平均每日去除量分别为0.47和0.37 mg·L-1, 试验后期去除较慢。皮尔逊相关性分析表明, 硝态氮日去除量、 总氮日去除量均与水体COD浓度呈极显著相关(相关性分别为0.96**、 0.93**)。因此, 沉水微宇宙对硝态氮的去除与氮源显著相关。
硝态氮单独施加时, 系统产气量先随时间逐渐升高, 第18天时达到最大值0.08 mg·L-1·d-1, 再逐渐下降, 第36天时达到较低值0.04 mg·L-1·d-1, 第48天达到次高峰0.07 mg·L-1·d-1后又开始下降。碳源与硝态氮共施时(图4b), 开始时系统气体产生量较低, 第6天为0.05 mg·L-1·d-1, 第12天开始产气量显著上升, 第30天达到最大值1.30 mg·L-1·d-1, 然后又急剧降至第42天的0.09 mg·L-1·d-1, 后期产气量较低。试验期间, 碳源硝态氮共施处理的总产气量为5.41 mg·L-1, 为硝态氮单独处理的9.18倍。外源施加硝态氮, 水体DO水平呈现先显著下降, 后保持平稳, 再上升的过程:第12天时DO水平较开始时下降了2.64 mg·L-1, 第66天时DO水平恢复至6.26 mg·L-1, 仍较初始时低(图4a)。 外源碳源硝态氮共施(图4b)显著降低水体溶解氧浓度, 试验第12天就降至厌氧水平(0.56 mg·L-1), 第12~36天基本保持不变, 第42天开始回升至接近初始水平(2.71 mg·L-1)。皮尔逊相关性分析结果表明, 碳源硝态氮处理的产气量与水体DO水体呈极显著负相关, 相关系数为-0.85**,这表明改变水体DO水平是碳源促进反硝化作用的机理之一。后期碳源消耗至较低浓度, 水体溶解氧浓度开始上升, 抑制了反硝化反应的进行。

图4 碳源对硝态氮净化效果的影响
硝态氮单独施加时, 植物对硝态氮的吸收量为(4.19±0.21)mg·g-1, 外源硝态氮与碳源共施处理的吸收量为(52.77±4.41)mg·g-1, 较前者增加了11.59倍。外源硝态氮和铵态氮分别单独施加时, 植物对氮源的吸收量分别为4.19和28.99 mg·g-1。与碳源共施时, 植物对硝态氮和铵态氮的吸收量分别为单独施加处理的12.59和1.82倍, 表明碳源对植物硝态氮的吸收的影响较其对铵态氮吸收的影响更显著。
2.3 碳源对苦草铵态氮硝态氮混合氮源净化效果的影响
由图5a可知, 施加混合氮源时, 铵态氮浓度和总氮浓度随时间逐渐下降, 总去除量分别为8.15和4.92 mg·L-1, 去除率分别为94.77%和40.27%。硝态氮浓度则随时间波动上升, 铵态氮硝化率(ANNR)和铵态氮净化率(ANRR)分别为32.75%和60.37%, 即硝化作用是铵态氮去除的主要途径之一。
当碳源与混合氮源共施时(图5b), 铵态氮浓度先随时间急剧下降, 降至较低浓度后(0.76 mg·L-1), 再平缓下降。铵态氮总去除量和去除率分别为8.06 mg·L-1和96.25%, 较混合氮源处理时不同, 碳源混合氮源共施时硝态氮浓度随时间呈波动下降的趋势, 试验结束时硝态氮浓度较初始硝态氮浓度下降了1.16 mg·L-1, 去除率为51.6%, 铵态氮硝化率(ANNR)为-14.43%。总氮浓度的去除趋势与铵态氮相似, 表现为前期急剧下降, 后期缓慢下降的趋势, 总去除量为9.86 mg·L-1, 去除率为90.84%。
由图6a可知, 混合氮源处理中, 水体溶解氧浓度先逐渐下降, 在第3天时降至最低值3.01 mg·L-1, 再波动回升。当碳源与混合氮源共施时, 水体溶解氧浓度急剧下降, 第1~4天时甚至达到厌氧水平, 第5天又开始波动上升, 但溶解氧浓度均低于混合氮源处理相同时间点的溶解氧水平。
由图6b可知, 混合氮源处理, 系统产气先增加, 第2天达到最大值0.08 mg·L-1·d-1, 之后又慢慢下降, 但产气量整体较低。外碳源与混合氮源共施时, 第1天时系统产气量很弱, 第2天却急剧增加, 达到最大值1.31 mg·L-1·d-1, 再急剧下降, 第6天时达到最小值0.02 mg·L-1·d-1, 此后产气量很微弱。第2~5天时铵态氮浓度和硝态氮浓度均下降, 且水体DO浓度达到厌氧水平。

图5 碳源对混合氮源净化效果的影响

图6 碳源混合氮源共施处理DO和产气量的变化规律
混合氮源处理时, 植物氮平均吸收量为(33.20±2.11)mg/g; 碳源与混合氮源共施处理时, 植物对氮源的吸收量较混合氮源处理高35.80 mg/g, 为(69.00±4.10)mg/g, 再次证明了碳源促进植物对氮源的吸收。
3 讨 论
3.1 促进植物对氮源的协同吸收
碳、 氮均为植物必需的大量元素, 是植物的重要组成成分。一定浓度范围内, 碳源和氮源共同处理时, 植物对氮源的吸收量大于氮源单独处理时的吸收量[13-14]。本研究结果与此类似, 碳源铵态氮共施处理植物对外源氮的净吸收总量为铵态氮单独施加处理的2.94倍; 碳源硝态氮共施处理则为硝态氮单独处理的12.59倍; 当以铵态氮和硝态氮为混合氮源, 碳源混合氮源共施处理的吸收量为混合氮源处理的2.08倍。吴文卫[15]通过对比碳源预处理和无碳源预处理对美人蕉氮吸收的影响研究发现, 碳源预处理显著促进美人蕉对硝态氮的吸收。水体中总氮浓度的变化也进一步印证了此观点。
研究还发现, 碳源共施能够促进系统对铵态氮、 硝态氮和混合氮源的去除, 表现为碳源铵态氮共施、 碳源硝态氮共施、 碳源混合氮源共施处理的TN去除量均显著高于各无碳源处理的去除量。当HRT为24 h, 硝态氮浓度为25~45 mg·L-1时, 投加25~50 mg的葡萄糖, 硝态氮去除率较不施葡萄糖处理增加了1倍以上[16],说明了碳源存在时促进植物对氮源的协同吸收是碳源促进微宇宙中氮源去除的重要途径之一。曹文平[17]以稻草作为植物生长的基质及缓释碳源的研究发现, 稻草释放的有机物符合一次线性方程, 并促进不同氮源按分数动力学(反应级数α<1)规律去除。
铵态氮和硝态氮是水体中主要的无机氮源。与单一氮源处理相比, 碳源硝态氮共施、 碳源铵态氮共施时植物对氮源的吸收量分别增加了11.59和1.94倍, 说明了碳源对硝态氮的植物协同吸收的影响更显著。这可能是因为植物对不同氮源的代谢机理不同[5], 且不同氮源对植物生理代谢的影响不同[18]。
3.2 碳源对氮源的形态转化及去除途径的影响
溶解氧是水体水质的重要指标, 水体溶解氧对水体的氧化还原性质、 水中物质的迁移转化、 水中动植物和微生物的活性具有重要影响。研究认为, 水生植物通过光合作用向水体分泌氧气, 显著影响水体的DO水平[11]。但铵态氮是还原性物质, 外源铵态氮在水中发生硝化作用会消耗水中的溶解氧, 降低水体溶解氧水平。研究发现, 当铵态氮浓度为7.97 mg/L时, 外源施加铵态氮使水体DO最大下降了4.30 mg/L(图2e);外源碳源与铵态氮共施时, 水体DO浓度最大下降了7.09 mg/L, 说明了外碳源的施入也消耗水体溶解氧, 加剧氮源对水体DO的影响。硝态氮的研究结果与此相似。
庄沐凡等[19]研究发现, 当以菖蒲残体为外加碳源时, 菖蒲残体添加量为0.5~1.5 mg·L-1时, 其腐解均使水体溶解氧降至厌氧水平。杨新平等[8]以柠檬酸盐、 乙酸盐、 葡萄糖为外碳源的摇瓶试验发现, 不同C/N处理下水体DO均快速下降至缺氧甚至厌氧水平, 随碳源的消耗, DO又开始恢复至好氧水平。在厌氧条件下, 硝态氮发生反硝化作用、 铵态氮发生厌氧氨氧化作用均产生N2、 N2O等气体, 因此, 氮化物气体的产量可以表征系统反硝化作用或厌氧氨氧化作用的程度。
碳源铵态氮共施(图3a)、 碳源硝态氮共施(图4b)以及碳源混合氮源共施(图6b)时系统产气量较氮源单独施加时显著增加, 说明了碳源共施促进硝态氮和亚硝态氮的还原以及铵态氮的厌氧氨氧化作用。这是因为碳源氮源共施显著降低水体溶解氧浓度, 使水体处于厌氧环境, 为系统厌氧氨氧化作用和反硝化作用的发生提供环境条件。与Boley 等[20]、 Shen等[21]、 Islam 等[22]的研究结果相似。因此,改变水体的性质, 改变氮源的转化途径, 使在厌氧条件下铵态氮发生厌氧氨氧化、 硝态氮发生反硝化也是碳源促进系统氮源去除的主要原因。铵态氮单独施加时, 铵态氮净化率ANRR仅为53.11%, 铵态氮硝化率ANNR为57.80%, 即部分铵态氮并没有彻底的从水体中去除, 而是发生了硝化作用, 以更稳定的硝态氮形式存在。但碳源与铵态氮共施时, 铵态氮净化率ANRR为93.35%, 铵态氮硝化率ANNR为1.71%, 说明了碳源共施促进了铵态氮的去除, 且铵态氮是彻底地从水体中去除。这是因为, 一方面碳源的共施改变了系统的好氧环境, 从而抑制了硝化作用的发生; 另一方面, 硝化菌为自养型微生物, 反硝化菌为异养型微生物, 碳源的存在会抑制硝化作用的进行, 而促进反硝化作用的进行[23]。忽略掉原始低浓度硝态氮的影响, 碳源铵态氮共施处理的铵态氮日平均去除速率为1.11 mg·L-1·d-1(试验前5 d), 较铵态氮单独处理增加了0.27 mg·L-1·d-1。
综上所述, 碳源的存在不仅改变了氮源的去除途径及不同去除途径的占比, 还改变了氮源的形态转化途径,说明了对水体水质的影响程度不同导致反硝化作用程度不同,以及植物对氮源的协同吸收程度不同是导致不同碳源对硝态氮的去除能力差异的原因。
3.3 反硝化作用
反硝化作用和厌氧氨氧化作用都是在厌氧环境下发生的[24]。碳源与铵态氮共施时, 试验第2~6天系统产气量较高(图 3a), 同时系统溶解氧浓度也达到了厌氧水平(图 2e), 因此, 此气体的增加可能是因为铵态氮发生了厌氧氨氧化作用产生的。由于第1天时系统处于好氧状态, 使系统中硝态氮浓度有了一定的积累, 因此, 此气体也可能是系统中积累的硝态氮发生了反硝化作用产生的。有研究发现, 厌氧氨氧化菌对外界条件较为敏感, 厌氧氨氧化作用启动较为困难[25]。首先, 厌氧氨氧化菌只有达到一定的种群密度才能产生明显的作用, 且其增殖速率很慢, 倍增时间为10~12 d[26], 由于初始时系统处于好氧状态, 因此厌氧氨氧化菌菌群密度不可能高, 在短时间内厌氧氨氧化菌不可能大量繁殖。其次, 厌氧氨氧化菌为厌氧自养型微生物, 以水中CO2和H2CO3为碳源, 而有机碳源会促进异养型微生物的生长, 从而抑制厌氧氨氧化菌的生长[27]。在本试验有(有机)碳源共存的情况下不利于厌氧氨氧化菌的生长繁殖,因此本试验条件下发生厌氧氨氧化作用的可能性较小。另外, 当铵态氮与硝态氮共存时, 植物对铵态氮吸收具有优先选择性[5], 在本试验铵态氮浓度较高的情况下, 植物对硝态氮的吸收很微弱, 此时硝态氮的减少是因为发生了反硝化作用。这是因为植物根系不仅为微生物提供栖息场所, 还通过光合作用在其根际形成好氧-厌氧的微域环境, 促进根系硝化反硝化作用的进行[28]。
综上所述, 碳源与铵态氮共施前期产生气体主要来自于早期积累的硝态氮的反硝化作用。研究还发现, 当碳源与硝态氮共施时, 在试验的前期也有一个产气量高峰(图 4b), 但产气量较碳源铵态氮共施(图 3a)时显著增加。这是因为碳源硝态氮处理的硝态氮浓度显著高于碳源铵态氮处理。在碳源与铵态氮共施处理中, 硝态氮浓度较开始时下降了。由于植物对铵态氮的优先选择性, 试验第2~5天时水体中铵态氮浓度还较高, 此时植物对硝态氮吸收较弱, 因此硝态氮浓度的下降可能是发生了反硝化作用。上述研究发现佐证了碳源铵态氮共施处理的产气量是发生了反硝化作用。
4 结 论
(1)碳源处理促进微宇宙对不同氮源的净化效果, 与无碳源处理相比, 有碳源处理的TN去除率分别提高了53.07%(铵态氮)、 39.66%(硝态氮)、 50.57%(混合氮源)。促进植物对铵态氮、 硝态氮、 混合氮源的吸收是外碳源促进微宇宙对不同氮源去除效果的主要机理之一。
(2)碳源对环境条件的改变从而影响氮源的迁移转化, 是其影响氮源去除效果的机理之一。外碳源加剧了氮源对水体DO的影响程度, 为铵态氮发生厌氧氨氧化和硝态氮发生反硝化作用提供环境条件。碳源铵态氮处理、 碳源硝态氮处理对DO的最大降低值分别较单一处理的最大降低值增加了2.79和4.08 mg·L-1。硝化作用是铵态氮主要的去除途径, 外氮源消耗水体的溶解氧, 抑制硝化作用的进行, 导致碳源铵态氮处理的铵态氮硝化率ANNR较铵态氮单独处理时低56.09%。
(3)硝态氮与铵态氮共存时, 在缺氧或厌氧条件下, 系统产生的气体主要是因为发生了反硝化作用, 即反应发生的难易程度:厌氧氨氧化作用难于反硝化作用。
猜你喜欢
河南师范大学学报(自然科学版)(2022年3期)2022-04-14
华北水利水电大学学报(自然科学版)(2020年6期)2021-01-29
农业环境科学学报(2020年10期)2020-11-11
中国酿造(2016年12期)2016-03-01
中国酿造(2014年9期)2014-03-11
植物营养与肥料学报(2012年1期)2012-10-26
植物营养与肥料学报(2011年5期)2011-11-06
植物营养与肥料学报(2011年4期)2011-10-26
植物营养与肥料学报(2011年6期)2011-10-24
植物营养与肥料学报(2010年3期)2010-11-16
