
松褐天牛幼虫肠道黏质沙雷氏菌培养条件与木质素降解功能*
2020-03-19 07:23傅慧静吴松青梁光红黄世国张飞萍
林业科学 2020年2期
傅慧静 胡 霞 吴松青 王 荣 梁光红 黄世国 张飞萍
(福建农林大学林学院 福州 350002)
木质纤维素是林木蛀干害虫幼虫阶段的主要营养来源,在该类昆虫的生长发育进程中起着重要作用(Jungetal.,2015;Petersonetal.,2015)。木质素是木质纤维素的主要成分(蒋挺大,2009;邱学青等,2013;De Gonzaloetal.,2016),属大分子物质,性状稳定,不溶于水和绝大多数溶剂(陈跃辉,2013),不能直接被昆虫肠壁细胞吸收运转。许多研究表明,昆虫肠道微生物在木质素生物降解过程中扮演着重要角色,其通过产生多种胞外酶推动木质素裂解反应,形成可被吸收的小分子物质(Buggetal.,2011;Wangetal.,2013;Lotfi,2014;Harithetal.,2014)。木质素过氧化物酶(lignin peroxidase,LiP)、锰过氧化物酶(manganese peroxidase,MnP)和漆酶(laccase,Lac)被认为在木质素降解过程中起重要作用,其活性反映了特定微生物的木质素降解能力(陈跃辉,2013)。
长期以来,针对昆虫肠道微生物产木质素降解酶的研究大多集中在真菌方面(喻云梅等,2005;彭木等,2014;Sanchez,2009),但昆虫肠道细菌也能够降解木质素。Delalibera 等(2005)在研究椴六点楔天牛(Saperdavestita)等昆虫的肠道微生物多样性时发现,源于椴六点楔天牛幼虫肠道中的细菌既能降解滤纸,也能降解羧甲基纤维素;Schloss等(2006)研究表明光肩星天牛(Anoplophoraglabripennis)肠道中的放线菌具有降解木质素类化合物的能力;Park(2007)等研究了9种天牛肠道细菌群落的多样性,并在松褐天牛(Monochamusalternatus)肠道内细菌降解木质纤维素的研究过程中分离得到具有木质素酶活性的变形菌门细菌(Gammaproteobacteria);袁俊超等(2015)以试材木质纤维为唯一碳源,从天牛幼虫的肠道微生物菌群中筛选出3株能够降解木质纤维的菌株,同时发现枯草芽孢杆菌(Bacillussubtilis)能协同烟曲霉(Aspergillusfunigatus)提高对木质素的降解效率;李成林(2015)从4种食木白蚁肠道中筛选出可转化利用木质素类化合物的共生细菌。
除此之外,细菌还存活于许多其他种类昆虫的肠道中,如棉铃虫(Helicoverpaarmigera)、家蚕(Bombyxmori)、舞毒蛾(Lymantriadispar)、小菜蛾(Plutellaxylostella)、美洲大蠊(Periplanetaamericana)、蜜蜂(Apiscerana)、桑天牛(Aprionagermari)、绿步甲(Carabussmaragdinus)和暗黑鳃金龟(Holotrichiaparallela)等(何正波等,2001;相辉等,2008;王振鹏,2009;黄胜威,2012;杨乐乐,2013;刘晓飞,2015;郭军等,2015;Crudenetal.,1984;Xiaetal.,2013;Huetal.,2017),其中一些肠道细菌已被证明能够产生1,4-β-内切葡聚糖酶、1,4-β-木聚糖酶、果胶酶、α-淀粉酶等,具有促进昆虫肠道消化吸收木质纤维素、木聚糖、果胶和淀粉等营养物质的功能(Dillonetal.,2004),但有关肠道细菌通过产木质素降解酶分解木质素的研究则较少。
沙雷氏菌(Serratiaspp.)隶属于变形细菌门(Proteobacteria)肠杆菌科(Enterobacteriaceae),通常存在于动植物、水、土壤和人体中。昆虫也是该类细菌的宿存对象,如宽须蚁蝗(Myrmeleotettixpolpalis)、白蚁(Reticulitermeshesperus)、菜青虫(Pierisrapae)、小菜蛾、南亚实蝇(Bactroceratau)、黑广肩步甲(Calosomamaximoviczi)、华山松大小蠹(Dendroctonusarmandi)和桑天牛等的肠道中均发现沙雷氏菌(金虹等,2005;姬小雪等,2009;李会平等,2012;夏晓峰等,2013;文竹等,2015;骆米娟等,2016;Thayer,1976)。有研究表明,该属的一些种类具有木质纤维素降解功能,如源于华山松大小蠹幼虫肠道中的沙雷氏菌能够分泌木质纤维素酶(胡霞,2014),Anand等(2010)发现液化沙雷氏菌(S.liquefaciens)具有分泌木质纤维素酶等多种胞外酶的能力,Perestelo等(1994)证实从堆肥中分离纯化出的黏质沙雷氏菌(S.marcescens)具有降解木质素的能力。然而有关昆虫肠道沙雷氏菌木质素降解功能的研究极少。
松褐天牛由于是松材线虫病(Bursaphelenchusxylophilus)的主要传播媒介而成为备受关注和危害极大的林木蛀干害虫(陈顺立等,1997;Zhaoetal.,2014),其食物的主要成分为木质纤维素。研究该虫对木质纤维素的吸收利用机制,有助于更好地理解其对寄主的适应性(王健敏等,2012)。笔者在前期研究该虫肠道微生物的过程中,采用以羧甲基纤维素钠作为唯一碳源培养,结合刚果红法筛选纤维素降解细菌,发现了154株分别隶属于变形菌门(Proteobacteria)、厚壁菌门(Firmicutes)和拟杆菌门(Bacteroidetes)8个属的纤维素降解细菌,其中黏质沙雷氏菌占比20.13%(胡霞等,2018),属于肠道优势细菌。为进一步探索该菌的功能及其作用机制,笔者研究其对木质纤维素主要成分木质素的降解功能,测定了其产木质素降解酶的情况,以及不同体外培养条件对其产优势木质素降解酶——木质素过氧化物酶的影响,以期为深入理解松褐天牛与该菌的协同作用提供基础依据。
1 材料与方法
1.1 试验材料
1.1.1 供试菌株 黏质沙雷氏菌HX-3菌株分离自松褐天牛幼虫肠道。松褐天牛幼虫来源于福建省福州市连江县琯头镇中榜水库周边的马尾松(Pinusmassoniana)林(26.150°N,119.593°E),并通过劈开马尾松木段的方法收集(胡霞等,2018),菌株低温保存于实验室。
1.1.2 主要培养基 SOC培养基(胡霞,2014):SOC 34 g;琼脂12 g;蒸馏水1 000 mL。硫酸盐木质素产酶培养基(陈跃辉,2013):KL(kraft lignin)3.0 g·L-1;(NH4)2SO42.0 g·L-1;K2HPO41.0 g·L-1;KH2PO41.0 g·L-1;MgSO40.2 g·L-1;CaCl20.1 g·L-1;FeSO40.05 g·L-1;MnSO40.02 g·L-1;pH7.0。该培养基中KL为唯一碳源。
1.2 试验方法
1.2.1 活化菌种 将经过纯培养得到的黏质沙雷氏菌单菌落接种在无菌SOC培养基内,置于30 ℃恒温、避光的条件下进行活化培养,并作为后续木质素降解试验的接种体。
1.2.2 发酵培养 将经过活化后的黏质沙雷氏菌接种于装有70 mL硫酸盐木质素液体培养基的250 mL规格锥形瓶内,200 r·min-1,30 ℃连续培养10天(Ahmad,2010),每天定时定量取样,并检测其木质素降解率与酶活性,3次重复。
1.2.3 黏质沙雷氏菌对木质素降解率的测定 以0、5、10、15、20、25、30、35、40、45、50 mg·L-1的质量浓度梯度配制木质素磺酸钙标准品溶液,在木质素特征吸收峰280 nm处,分别检测不同质量浓度下各标准品的OD值(每个质量浓度重复 3 次),再以OD值为纵坐标,各质量浓度为横坐标,绘制标准曲线,并建立回归分析直线方程(图1)。每天于超净工作台内,通过移液枪所吸取的液体培养基样品,以12 000 r·min-1转速离心10 min,取其上清液,测定各样品在280 nm处的OD值(陈跃辉,2013;乔乔,2013)。最终依据标准曲线方程求解出目标菌株的木质素质量浓度,再代入下式计算出木质素的降解率。
木质素降解率=
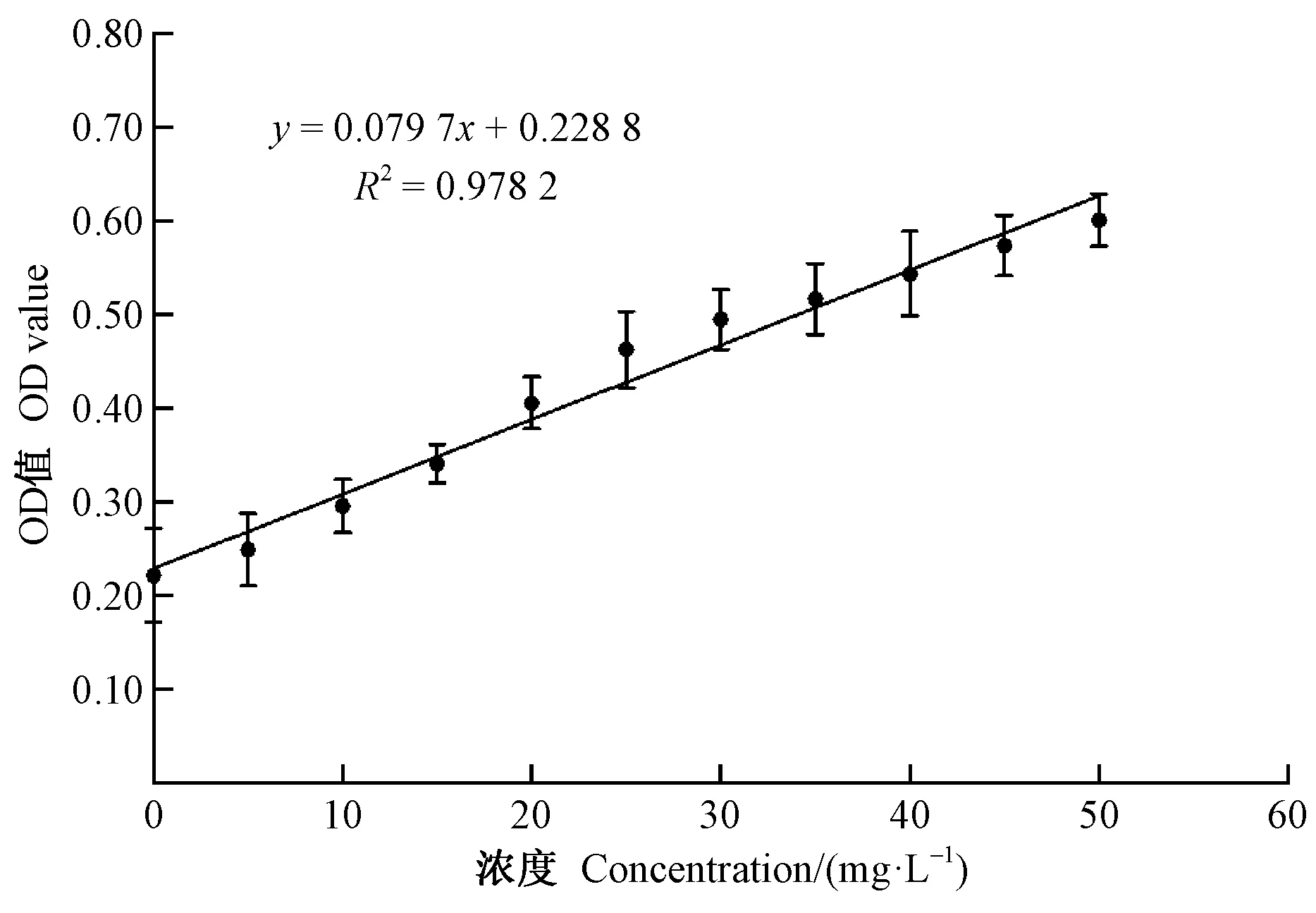
图1 木质素标准曲线Fig.1 Standard curve of lignin
1.2.4 粗酶液的制备 取培养后的菌液,以4 ℃、10 000 r·min-1,离心5 min,取其上清液即为粗酶液。
1.2.5 黏质沙雷氏菌产木质素降解酶活性的测定 按照1.2.2的方法培养菌液发酵产酶,并于培养的第1—10天分别每日定时定量取样以制备粗酶液。同时,以未接菌种的相同培养基作为空白对照(3次重复)。酶活力为酶促反应在单位时间内释放出1 μmol产物所需的酶量,定义为1个酶活单位(U·L-1)(Kapichetal.,2004)。
木质素过氧化物酶(LiP)活性检测:依据相关学者的研究,选用藜芦醇法(Mingetal.,1988)。具体为:首先,借助酶标仪设定30 ℃恒温反应条件,利用移液枪分别吸取2 mmol·L-1VA 30 μL、50 mmol·L-1酒石酸钠缓冲液(pH 2.5)139和30 μL的粗酶液于酶标板(CORNING,3590)上,随后通过1 μL 0.4 mmol·L-1H2O2启动体系的反应,并于310 nm的波长处读取、记录1 min内体系OD值的变化。
锰过氧化物酶(MnP)活性检测:参照Wariish等(1992)的相关研究,选用二价锰氧化法。首先,借助酶标仪设定30 ℃恒温反应条件,利用移液枪分别吸取1 mmol·L-1MnSO430 μL、50 mmol·L-1丙二酸钠缓冲液(pH 4.5)139 μL和粗酶液30 μL于酶标板(CORNING,3590)上,随后用1 μL 0.1 mmol·L-1H2O2启动体系的反应,并于270 nm的波长处读取、记录1 min内体系OD值的变化。
漆酶(Lac)活性检测:参考Wolfenden等(1982)的研究,采用ABTS法。具体为:首先,借助酶标仪设定30 ℃恒温反应条件,利用移液枪分别吸取1 mmol·L-1ABTS 30 μL和100 mmol·L-1的醋酸钠缓冲液(pH5)140 μL于酶标板(CORNING,3590)上,后续用粗酶液30 μL启动体系的反应,并于420 nm的波长处读取、记录1 min内体系OD值的变化。
1.2.6 不同培养条件对黏质沙雷氏菌产Lip活性的影响 结合1.2.3和1.2.5研究结果,选择活性最高的优势木质素降解酶——LiP为对象,并选取产酶高峰期第4天的17:00为取样时间,测定不同培养条件对黏质沙雷氏菌产酶活性的影响,3次重复。
不同木质素质量浓度的影响:以硫酸盐木质素液体产酶培养基为基础,分别设置9组不同浓度(质量浓度)梯度的木质素,具体为0、1.0、2.0、3.0、4.0、5.0、10.0、15.0、20.0 g·L-1。其余条件和方法同木质素过氧化物酶(LiP)活性的测定。
不同pH值的影响:以硫酸盐木质素液体产酶培养基为基础,利用pH 计(上海奥豪斯仪器有限公司)通过1 mol·L-1HCl和1 mol·L-1NaOH调节并设置7组不同的pH值处理,分别为pH4、5、6、7、8、9、10。其余条件和方法同木质素过氧化物酶(LiP)活性的测定。
不同氮源种类的影响:以硫酸盐木质素液体产酶培养基为基础,分别设置2组不同的氮源种类:有机氮源、无机氮源,有机氮源具体为蛋白胨(peptone)、酵母膏(yeast extracts)、干酪素(casein);无机氮源具体为(NH4)2SO4、NH4NO3、NH4Cl、尿素[CO(NH2)2]。其余条件和方法同木质素过氧化物酶(LiP)活性的测定。
不同酵母膏(氮源)浓度的影响:基于不同氮源种类影响的研究结果显示酵母膏产LiP的活性最强,故而选取酵母膏为氮源,进一步检测不同氮源浓度对LiP酶活性的影响。同样以硫酸盐木质素液体产酶培养基为基础,利用电子分析天平称取酵母膏以设置7组不同的浓度(质量浓度)处理,具体为0、1.0、2.0、3.0、4.0、5.0、10.0 g·L-1。其余条件和方法同木质素过氧化物酶(LiP)活性的测定。
不同金属离子浓度的影响:以硫酸盐木质素液体产酶培养基为基础,通过电子分析天平称取金属化合物以分别设置5组不同的金属离子浓度(质量浓度)梯度处理,具体为Mg2+、Ca2+、Fe2+、Mn2+、K+。其中,MgSO4分别为0、0.20、0.40、0.60、0.80、1.00 g·L-1;CaCl2分别为0、0.10、0.20、0.30、0.40、0.50 g·L-1;FeSO4分别为0、0.05、0.10、0.15、0.20 g·L-1;MnSO4分别为0、0.02、0.04、0.06、0.08、0.10 g·L-1;K2HPO4和KH2PO4的分别为0、1.00、2.00、3.00、4.00、5.00 g·L-1。其余条件和方法同木质素过氧化物酶(LiP)活性的测定。
1.3 数据处理
采用SPSS 22.0软件,利用单因素方差分析对试验所得数据进行统计分析;通过Duncan检验作不同处理间的差异显著性分析(P<0.05);应用PRISM 5.0与ORIGIN 9.0软件绘图。
2 结果与分析
2.1 黏质沙雷氏菌对木质素的降解功能
通过10天的液体发酵培养,黏质沙雷氏菌对硫酸盐木质素的降解情况见图2。由图可知,在木质素培养基中培养的前4天,其木质素降解率随时间的延长而增加,只是前2天的累计降解率仅为10.90%;随后在第4天达到降解高峰,单日降解率高达15.16%,并于维持3天的稳定期后,累计降解率达到66.23%。第7天降解率开始锐减,且随时间的推移降解率逐步降低,至第10天仅为3.98%,但累计降解率达到94.12%。上述结果说明,黏质沙雷氏菌对木质素具有显著的降解能力。
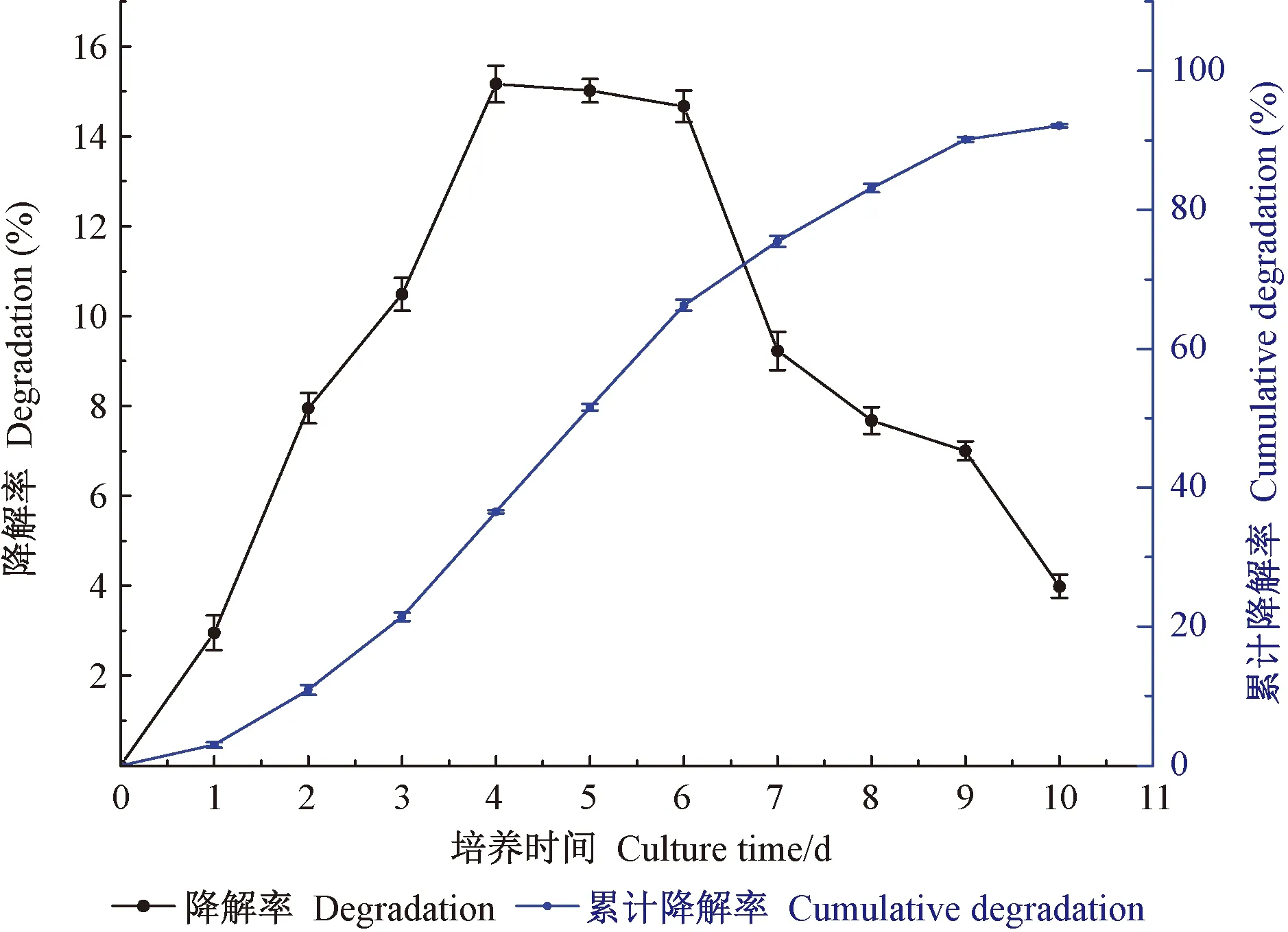
图2 松褐天牛幼虫肠道黏质沙雷氏菌木质素降解率Fig.2 Rate on degrading lignin by S.marcescens in the larval gut of M.alternatus
2.2 黏质沙雷氏菌产木质素降解酶特性
黏质沙雷氏菌在不同培养时期产木质素降解酶的情况见图3。该菌在各阶段产Lip酶的活性均最高,然后依次是MnP和Lac。对于Lip酶活性,初期伴随着培养时间的推移而逐渐上升,于第4天达到峰值(214.67 U·L-1),并在第5天开始呈现下降趋势,至第10天其活性仅为36.26 U·L-1;对于MnP酶活性,初期2天的酶活性处于快速增长的阶段,随后3天增长趋势则相对缓慢,但仍维持在较高的活性状态,最高峰出现在第5天,为145.47 U·L-1,而后开始逐步下降。Lac酶在整个培养期间的活性始终处于较低水平,且仅在培养的第3天出现1个小高峰,活性也只达到16.88 U·L-1。结合图2和图3可知,黏质沙雷氏菌对木质素的降解率与其产LiP、MnP2 种酶活性的日变化趋势相近,二者均在前期随着培养时间延长而逐步升高,并均在第4—5天期间出现活性高峰期,随后均逐步下降直至第10天。这说明黏质沙雷氏菌主要通过产LiP、MnP2 种酶而实现其木质素降解功能。
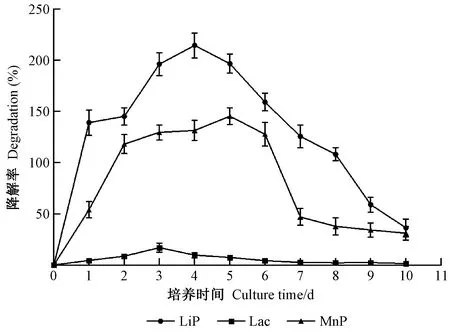
图3 不同培养时间松褐天牛幼虫肠道黏质沙雷氏菌产酶曲线Fig.3 Curves of enzyme production by S.marcescens in the larval gut of M.alternatus with different culture time
2.3 不同培养条件对黏质沙雷氏菌产Lip活性的影响
2.3.1 不同木质素浓度的影响 不同木质素浓度下LiP活性见图4。黏质沙雷氏菌产LiP的活性受木质素浓度影响显著(F=530.59,df=8,P<0.001)。当质量浓度处于0~3 g·L-1之间时,LiP的活性随木质素浓度的增加而显著提高,此后,伴随浓度的进一步增加,LiP的活性反而逐渐转弱。当木质素质量浓度为3 g·L-1时,酶活性达到峰值,且显著大于其他浓度处理。但当木质素质量浓度处于20 g·L-1时,酶活性却明显弱于除对照外的其他浓度处理。
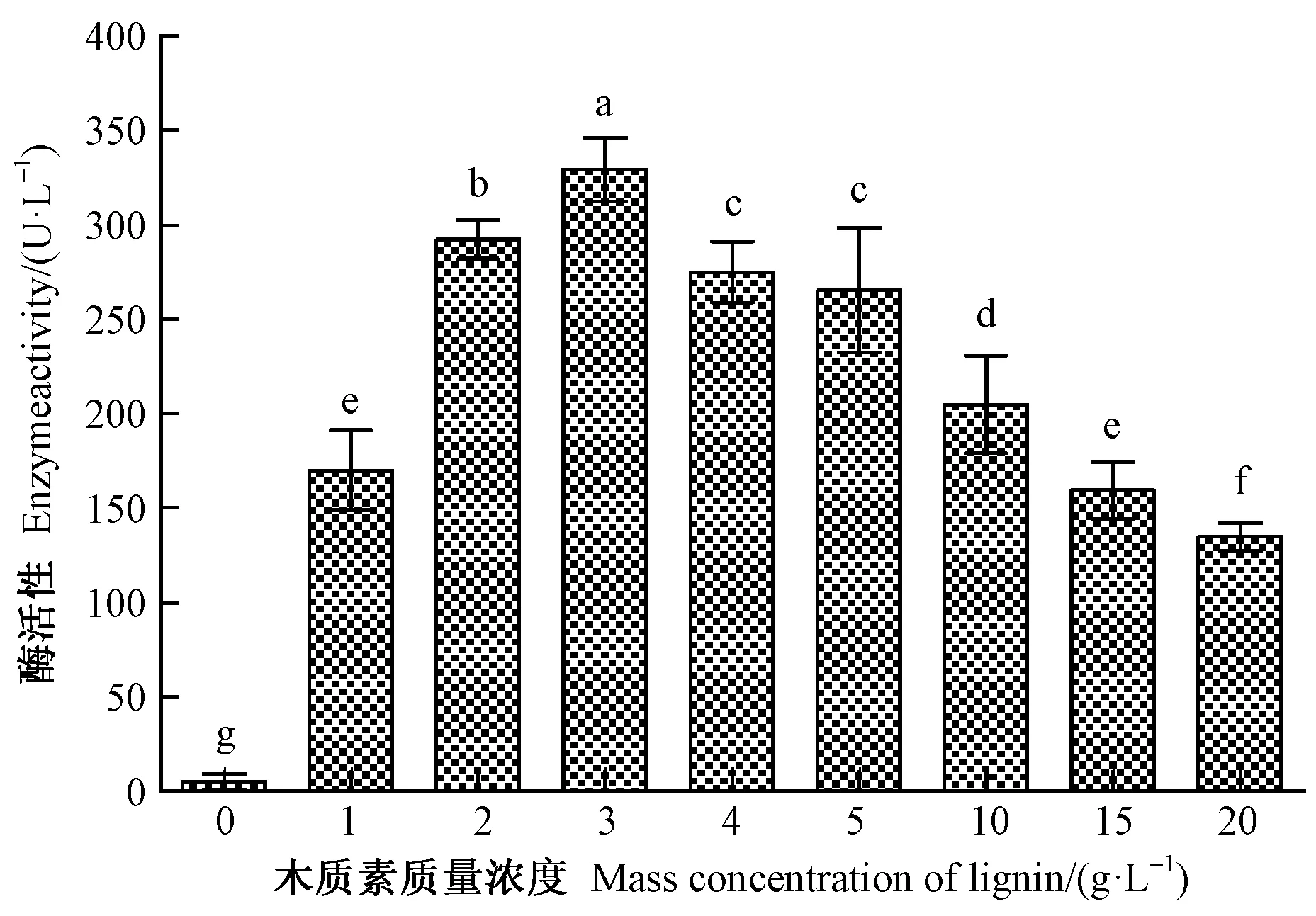
图4 不同木质素浓度下黏质沙雷氏菌产木质素过氧化物酶活性Fig.4 LiP activity from S.marcescens cultured with different conditions of lignin concentration图中垂直线表示标准误差,不同小写字母表示差异显著性(P<0.05)。下同。The vertical line represents the standard error,different lowercase letters indicate significant difference at 0.05 level.The same below.
2.3.2 不同pH值的影响 不同pH值对LiP活性的影响见图5。不同pH值对黏质沙雷氏菌产LiP的活性存在极显著差异(F=596.67,df=6,P<0.001)。处于偏酸性条件下的LiP活性显著超出偏碱性条件下的酶活性,当pH值为5时的LiP活性最大(344.63 U·L-1),且明显比其他pH值处理领先;此外,而在pH≥8后的3个处理中,LiP活性均显著低于其他pH值处理,说明碱性环境不利于黏质沙雷氏菌产LiP。
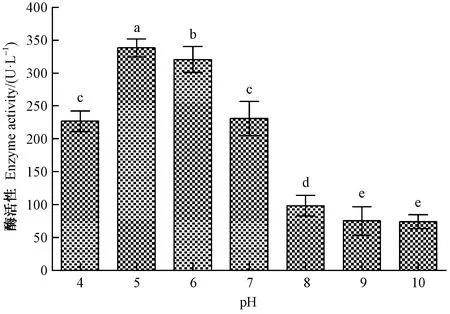
图5 不同pH下黏质沙雷氏菌产木质素过氧化物酶活性Fig.5 LiP activity from S.marcescens cultured with different pH conditions
2.3.3 不同氮源的影响 由图6可知不同氮源培养条件下LiP的活性情况:在有机氮源组中,蛋白胨、干酪素与酵母膏之间对黏质沙雷氏菌产LiP活性的影响具有显著差异(F=169.29,df=2,P<0.05),当氮源为酵母膏时,LiP活性最强(1 080.23 U·L-1);氮源为干酪素时,LiP活性最弱(400.74 U·L-1)。在无机氮源组中,(NH4)2SO4、NH4NO3、NH4Cl与CO(NH2)2之间对黏质沙雷氏菌产LiP活性的影响也存在显著性差异(F=212.17,df=3,P<0.05),其中氮源为CO(NH2)2时的LiP活性最强(374.07 U·L-1),而氮源为 (NH4)2SO4时的LiP活性最弱(207.20 U·L-1)。从整体看来,有机氮源组的酶活性显然强于无机氮源组,进一步表明有机氮源更适于黏质沙雷氏菌产LiP。
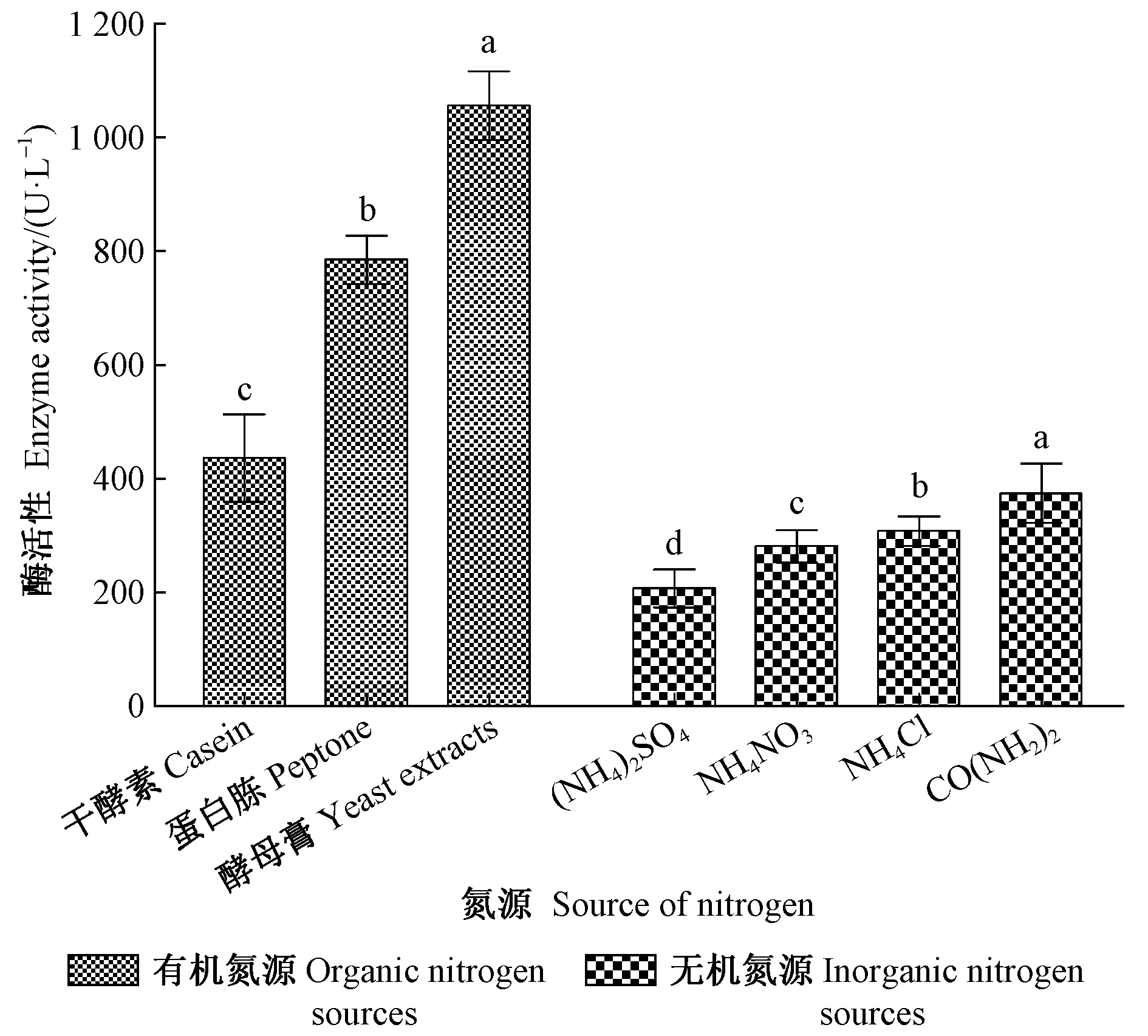
图6 不同氮源下黏质沙雷氏菌产木质素过氧化物酶活性Fig.6 LiP activity from S.marcescens cultured with different sources of nitrogen
2.3.4 不同酵母膏(氮源)浓度的影响 不同酵母膏浓度对LiP活性的影响见图7。酵母膏浓度对LiP活性具有显著影响(F=437.83,df=6,P<0.05)。在1~5 g·L-1范围内,LiP活性随酵母膏浓度增加而逐渐增大,尤其质量浓度为5 g· L-1的LiP活性最高,达633.63 U·L-1,显著高于其他各浓度处理;质量浓度为1 g·L-1的LiP活性最低(237.84 U·L-1)。
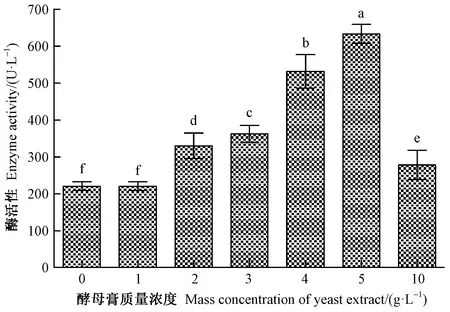
图7 不同酵母膏质量浓度下黏质沙雷氏菌产木质素过氧化物酶活性Fig.7 LiP activity from S.marcescens cultured with different mass concentration of yeast extract
2.3.5 不同浓度金属离子的影响 黏质沙雷氏菌在各金属离子不同浓度培养条件下产LiP活性如图8。从图可知,Mg2+浓度对该菌产LiP活性的影响达到显著水平(F=399.94,df=5,P<0.05),质量浓度为0.20 g·L-1时的活性最高,达221.31 U·L-1,显著高于其他各浓度处理;但当质量浓度处于1.00 g·L-1时的活性最低,仅为64.20 U·L-1,显著小于其他处理。
对于Ca2+,不同浓度之间差异显著(F=206.63,df=5,P<0.05)。当质量浓度为0~0.40 g·L-1时,随着Ca2+浓度增加,LiP活性逐渐上升,在质量浓度为0.40 g·L-1时的LiP活性最高(401.35 U·L-1),显著高于其他各浓度处理。
对于Fe2+,各浓度间也存在显著性差异(F=288.14,df=4,P<0.05),当质量浓度处于0.15 g·L-1时,酶活性最强(448.04 U·L-1),且明显强于其他各处理。而当质量浓度大于0.15 g·L-1时,酶活性则显著降低,且当质量浓度为0.20 g·L-1时的活性最低,仅为200.19 U·L-1,显著低于其他各处理。
对于Mn2+,不同浓度对LiP活性也具有显著影响(F=238.61,df=5,P<0.05),当质量浓度处于0.04 g·L-1时的酶活性最强(376.73 U·L-1),且显著强于其他各处理。但当Mn2+质量浓度大于0.04 g·L-1后,酶活性却迅速降低,并在质量浓度处于0.10 g·L-1时的酶活性仅为171.95 U·L-1。
对于K+,各浓度之间存在显著的差异(F=770.84,df=5,P<0.05),当质量浓度为0 g·L-1时,LiP活性最高(459.15 U·L-1),显著高于其他处理,并在质量浓度0~5.00 g·L-1范围内,LiP活性随着浓度增加而逐渐降低。
3 讨论
黏质沙雷氏菌在硫酸盐木质素液体培养基中培养10天,对木质素的累计降解率高达94.12%。这一结果证实来源于松褐天牛幼虫肠道中的黏质沙雷氏菌具备降解木质素的功能,且该菌培养期间处于第4天时对木质素的降解率最高(15.16%),这可能因为培养初期为菌体细胞急剧增长的对数期,该时期的细菌又恰恰是细胞生长代谢活动最为旺盛的时期,而kraft木质素正好作为菌株生长和代谢的唯一碳源被消耗利用,并为其后续的生长、发育、繁殖等生命活动提供能量。
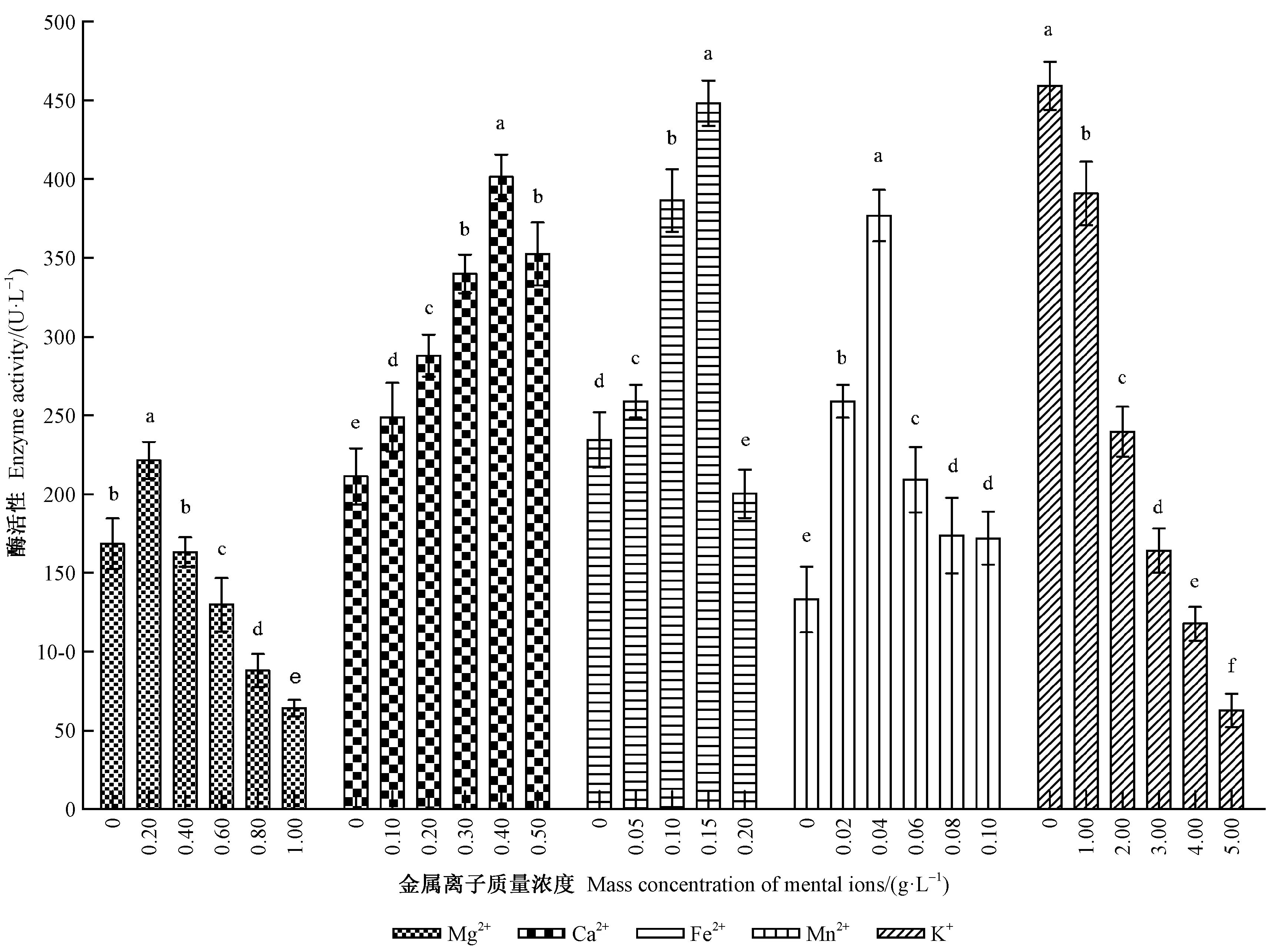
图8 不同Mg2+、Ca2+、Fe2+、Mn2+、K+离子质量浓度下黏质沙雷氏菌产木质素过氧化物酶活性Fig.8 LiP activity from S.marcescens cultured with different concentrations of Mg2+,Ca2+,Fe2+,Mn2+,K+
作为高分子聚合物的木质素直接进入单细胞细菌体内存在一定的难度,故不少细菌往往会选择通过分泌一系列胞外氧化酶这一与真菌相仿的方式完成对木质素的生物降解(Ramachandraetal.,1988;Buggetal.,2011;Charles,2013;Harithetal.,2014)。引起木质素降解的实质是由于在C—C键裂解的最初阶段能够致使木质素脱甲基形成含苯氧基自由离子,而这一氧化过程需要多种酶的协同作用(Deetal.,2016)。目前,国内外多方面的研究资料表明主要有3种酶参与木质素的降解,分别为LiP、MnP和Lac(Wangetal.,2013;Harithetal.,2014)。因而,这3种酶活性的强弱能在一定程度上衡量微生物降解木质素的能力。
本研究说明黏质沙雷氏菌能够通过分泌LiP、MnP和Lac实现对木质素的生物降解,其中LiP与MnP具有较高的活性,但Lac的活性在整个培养过程中始终处于较低的水平,仅在培养的第3天出现小高峰(16.88 U·L-1),明显低于LiP、MnP。而前两者分别在培养第4天与第5天的活性达到峰值(LiP:214.67 U·L-1;MnP:145.47 U·L-1),且其随时间的变化趋势与该菌对木质素的降解率相吻合,这既说明黏质沙雷氏菌可通过分泌LiP与MnP 2种胞外酶实现其木质素降解功能,同时也证明并非每种菌都依赖这3种酶降解木质素的说法(王靖等,2008)。
对特定种类的微生物,其木质素降解功能与其宿存的微生态环境如微量元素、环境pH值、诱导剂、氮源种类及其浓度等密切相关(高大文等,2005;乔乔,2013;徐海涛,2014;Mikaelyanetal.,2014)。本研究发现,木质素浓度、pH值、氮源种类、氮源浓度与金属离子及其浓度对黏质沙雷氏菌产LiP活性均具有显著影响,进而影响着其对木质素的降解功能。当木质素处于低浓度时,细菌降解木质素的反应速率与木质素浓度之间基本呈正相关。然而,随着木质素浓度增大,反应速率却不再上升(金剑等,2010),本研究中也存在类似情况,即该菌产LiP的活性并不伴随木质素浓度增加而增强,由此可见木质素浓度与LiP活性之间并不是呈现出浅显的线性关系,浓度越大并非对酶活性的提高越有优势。究其原因很可能是因木质素浓度增加而导致降解环境随之变得黏稠,阻碍体系内营养物质的流通与气体的流动,由此造成高浓度木质素条件下细菌的生长效果反而不如低浓度时理想,从而表现为高浓度时的酶活性不强。
pH值是影响微生物产木质素降解酶的重要因素,如节杆菌属(Arthrobactersp.)产木质素过氧化物酶的最佳pH值为7(张庆芳等,2014),斜卧青霉菌P6(PenicilliumdocumbensP6)产LiP的最适pH为4.0(杨金水等,2004),而本研究中发现黏质沙雷氏菌处于偏酸性环境中LiP活性较强,且最佳pH值为5,这同学者Ahmad(2010)所提出的观点“微生物产LiP在低pH时效果更佳”不谋而合。已有多方面的研究发现,木质素降解酶的合成与活性的强弱受培养基环境的pH影响较深。无论过高还是过低的pH值,均会显著影响酶的活性(张庆芳等,2014),这在本研究的分析中也得到了进一步的证实。
针对不同氮源对黄曲霉LDY(AspergillusflavusLDY)降解木质素影响的研究表明,有机氮源的降解效果均劣于无机氮源(乔乔,2013)。本研究发现有机氮源比无机氮源更适于黏质沙雷氏菌产LiP,这说明不同微生物对不同种类氮源的利用能力具有差异。此外,秦岭细粘束孢(Leptographiumqinlingensis)对寄主华山松金属离子的营养具有高度选择性(蒲晓娟等,2008)。而在本研究的分析中也发现不同金属离子浓度对于黏质沙雷氏菌产Lip活性具有显著影响,对Mg2+、Mn2+与K+离子来说,处于高浓度时LiP活性显然不如低浓度时的强,表现为抑制作用;而对于Ca2+与Fe2+,则表现为高浓度促进作用。以上结果说明黏质沙雷氏菌产Lip时对于不同金属离子的营养需求不同,进而可能进一步表现出对寄主树木营养的选择性。
本文为深入了解松褐天牛对寄主植物的营养适应机制提供了基础依据。然而,关于该虫肠道内的其他微生态环境条件、该菌在寄主肠道内产其他种类酶的情况以及与其他种类微生物的协同作用等,还有待于进一步地探索。
4 结论
黏质沙雷氏菌具有较强的木质素降解能力,可通过产生木质素过氧化物酶和锰过氧化物酶实现其对木质素的降解功能;培养基中木质素浓度、pH值、氮源种类及其浓度、金属离子及其浓度等对该菌产木质素过氧化物酶的活性均有显著影响。
猜你喜欢
河北科技师范学院学报(2022年2期)2022-08-26
上海包装(2019年8期)2019-11-11
天津造纸(2016年1期)2017-01-15
中国塑料(2016年3期)2016-06-15
中国酿造(2016年12期)2016-03-01
中国造纸学报(2015年1期)2015-12-16
橡胶工业(2015年4期)2015-07-29
橡胶工业(2015年3期)2015-07-29
中国塑料(2014年4期)2014-10-17
中国酿造(2014年9期)2014-03-11
