
成年斑胸草雀HVCX神经元电生理特性的侧别和性别差异
2020-03-23 05:14李东风
辽宁师范大学学报(自然科学版) 2020年1期
李东风, 冯 凯
(华南师范大学 生命科学学院,广东 广州 510631)
斑胸草雀是研究语言发声的模式动物,其高级发声中枢HVC(higher vocal center)与人类大脑中的布洛卡区同源,是鸣禽脑内控制鸣曲学习前端脑通路(anterior forebrain pathway, AFP)和控制鸣曲发声运动通路(vocal motor pathway, VMP )的共同起点.HVC核团中的HVCX神经元和HVCRA神经元通过神经投射分别控制AFP通路与VMP通路的X区与弓状皮质栎核(robust nucleus of the arcopallium, RA)[1].此外,HVC中还含有一类中间神经元HVCINT,在HVC核团内部分别对上述两类神经元产生抑制作用[2-4].HVCX神经元不仅将听觉信息传递给APF通路,而且APF通路的听觉应答也受HVCX神经元的影响.AFP通路主要参与鸣曲的学习过程,VMP通路主要负责鸣曲的发声行为.鸣曲的习得可以帮助斑胸草雀求偶、防御以及维护领地等社会性行为.
成年雄性斑胸草雀擅长鸣唱,而雌性不会鸣唱.在雌性斑胸草雀皮下埋植睾酮后,可显著增加HVC核团的体积和神经元数量,提示雄激素在鸟类鸣唱系统中发挥着不可替代的作用[5].此外,本实验室前期工作表明,损毁左侧HVC核团,对长鸣和鸣曲的频域和声强特征都无显著影响,而损毁右侧HVC后,则会导致鸣曲特征的显著性改变,包括鸣曲的振幅、平均频率和峰频率的显著性降低等[6].然而雌雄两性个体的HVCX神经元电生理特性差异尚不清晰,且损毁HVC是破坏整个脑区的功能,并不能反映单个神经元电生理特性的侧别差异.因此,基于前期工作的基础,探究HVC核团神经元电生理特性是否具有侧别差异和性别差异为本研究目的.
1 实验材料与方法
1.1 实验动物
成年雌雄斑胸草雀(zebra finch,Taeniopygiaguttata)(>90 d).体重9~15 g,购于广州花地湾花鸟虫鱼市场.
1.2 试剂药品
人工脑脊液ACSF(mmol/L):NaCl 125,NaHCO325,NaH2PO4·H2O 1.27,KCl 2.5,MgSO4·7H2O 1.2,CaCl22.0,Glucose 25;切片液(mmol/L):Sucrose 248,KCl 5,NaHCO328,Glucose 10,MgSO4·7H2O 1.3,NaH2PO4·H2O 1.26;电极内液(mmol/L):KMeSO4120,NaCl 5,HEPES 10,EGTA 2,Mg-ATP 2,Na-GTP 0.3,pH 7.2.
1.3 主要仪器
膜片钳放大器,MultiClamp 700B(Axon Instrument, USA);模/数转换器,DIGIDATA 1440A(Axon Instrument, USA),正置显微镜,BX51WI(Olympus, Japan),微电极拉制仪,P-97(Sutter Ins, USA),振动切片机NVSIMI(WPI, USA),分析天平等.
1.4 脑片制备
10%(质量分数)水合氯醛(0.05 mL/g)进行鸟胸大肌注射,待麻醉后快速断头取脑,置于切片液的冰水混合物中冷却30~60 s,用振动切片机在冰水混合的切片液中将大脑进行矢状切,切取含有HVC核团的250 μm厚的脑片后,将脑片移至温度为37 ℃且已通混合气(95%O2+5%CO2)30 min的孵育槽中孵育1 h,之后将温度调整为35 ℃,孵育0.5 h后进行膜片钳实验.
1.5 膜片钳全细胞记录
1.5.1 电极制备及内液灌注 用微电极拉制仪将硬质玻璃毛坯管(1.5 mm outer diameter, 1.17 mm inner diameter,length 8 cm, Sutter Ins, USA)拉制成微电极.全细胞记录电极尖端的阻抗为3~6 MΩ.
1.5.2 HVC神经元观察及记录 在显微镜下观察置于记录槽内的脑片, 寻找合适的HVC神经元.在电极接触神经元后对细胞给予负压进行封接,形成高达GΩ电阻的细胞吸附式,进一步给予负压进行破膜,形成全细胞记录模式.在电流钳模式下记录HVC投射神经元的诱发电位及阈值和后超极化幅值等各项电生理指标.
1.6 数据采集及统计分析
用pClampex 10.3软件采样系统获得信号后,根据HVC各类神经元的发放特性,将记录到的神经元进行分类.运用Origin 8.0软件双尾t-test检验方法对记录到的HVCX神经元的电生理特性指标进行统计与分析.统计指标如下:
(1)静息电位(resting potential,RP):指细胞膜未受刺激时,存在于细胞膜内外两侧的外正内负的电位差.
(2)动作电位潜伏期(evoked action potential latency):诱发刺激开始至第一个诱发动作电位起始所经历的时间.
(3)后超极化达峰值时间(AHP time to peak):动作电位下降相降至阈值时起算,直至AHP达最低点时所经历的时间.
(4)动作电位发放频率(spike rate):对500 ms、100 pA刺激下诱发的动作电位发放个数进行比较.
2 实验结果
2.1 HVC神经元的分类
根据HVC核团中3类神经元不同的发放特性,将记录到的神经元进行分类[7-8].HVCX神经元具有内向整流特性,在受到超极化电流刺激时具有明显的膜电位凹陷(sag)现象.除此之外,在受到持续地去极化刺激时,发放频率较高且存在适应性.中间神经元HVCINT具有强烈的自发放活动,受到超极化刺激时,同样存在sag现象,且比HVCX神经元更明显.此外,在超极化刺激结束后有一个反弹放电现象,HVCINT接受持续去极化刺激时会爆发一种发放持续、规则的紧张型高频放电模式.HVCRA神经元的静息电位高于另外两类神经元,受到超极化刺激时不存在sag现象,刺激结束后也没有反弹放电现象.受到去极化刺激时,只发放少数几个甚至是单个的动作电位(图1).研究表明,在鸣禽发声或听到多种鸣声刺激时,HVCX神经元都有相同的发放模式,显示出了精确的感觉运动一致性[9].故本实验以HVCX神经元为目标神经元,探究HVCX神经元的侧别差异和性别差异.本实验通过观察在电流钳下给予神经元500 ms、100 pA的去极化刺激和500 ms、-200~0 pA、步阶+20 pA的超极化刺激下的发放特性来鉴定神经元类型.
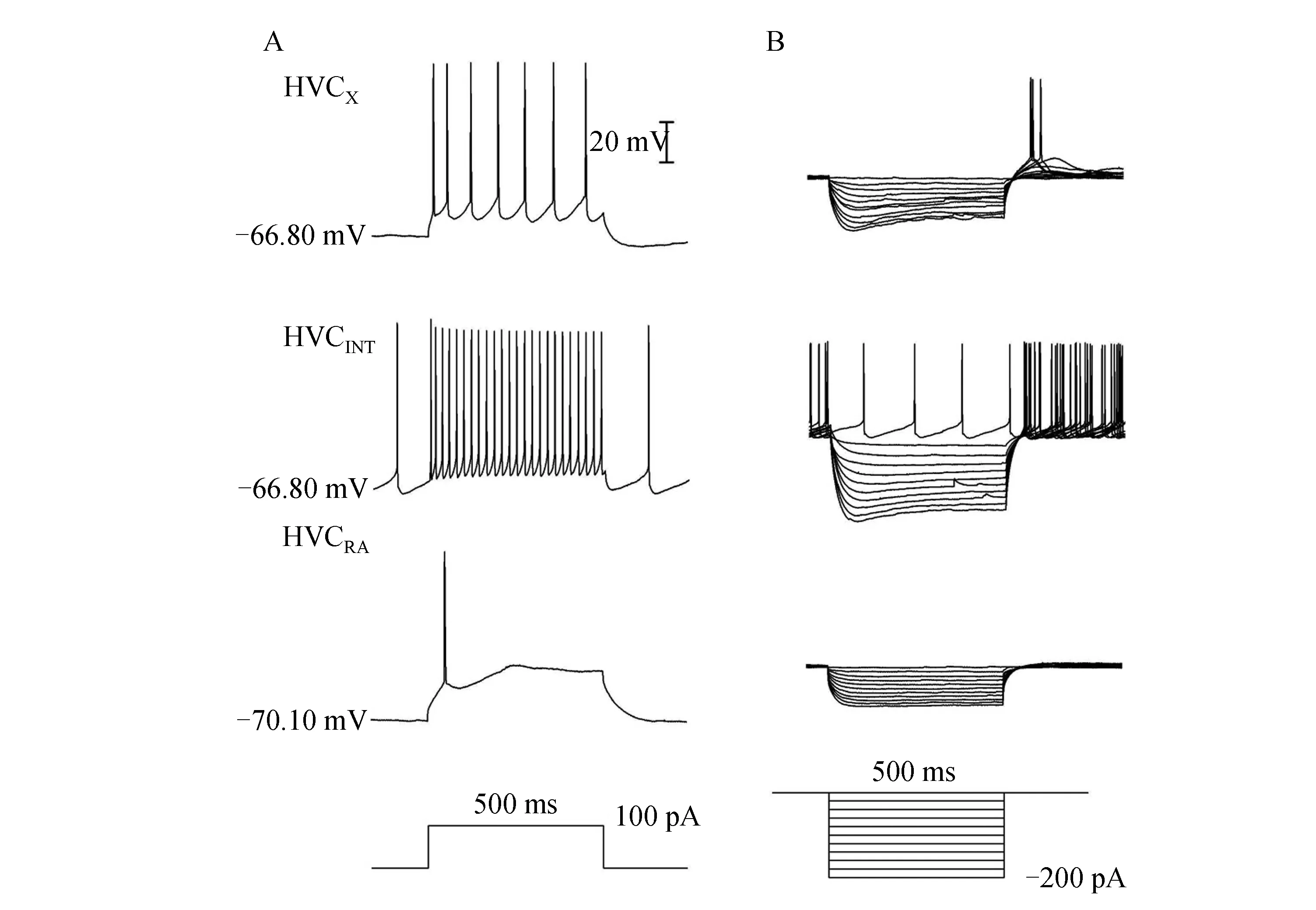
图1 HVC核团中3类神经元发放特性Fig.1 The firing properties of 3 types of neurons in HVC nucleus
2.2 雌鸟左右脑HVCX神经元电生理特性的侧别比较
对雌鸟左右脑HVCX神经元施加5 ms、300 pA电流刺激,将记录到的电生理指标进行统计.结果显示,雌鸟右脑的静息电位值((-61.63±3.77)mV,n=5)显著低于雌鸟左脑((-66.57±9.51)mV,n=14,P<0.05,图2B).但是,动作电位潜伏期(图2C)和后超极化达峰值时间(图2D)无显著性差异.

*P<0.05,差异且有显著性;**P<0.01,差异极显著图2 雌鸟左右脑HVCX神经元电生理特性比较Fig.2 The lateral comparison of electrophysiological properties in adult female zebra finches
2.3 雄鸟左右脑HVCX神经元电生理特性的侧别比较
对雄鸟左右脑HVCX神经元施加5 ms、300 pA电流刺激,将记录到的电生理指标进行统计.结果显示,右脑HVCX神经元的动作电位潜伏期((2.58±0.37) ms,n=5)低于左脑((3.89±0.48) ms,n=12),且差异极显著(P<0.01,图3C).右脑的后超极化达峰值时间((27.3±7.11) ms,n=5)低于左脑((41.83±6.90) ms,n=6),且差异极显著(P<0.01,图3D).而静息电位(图3B)无显著性差异.说明右脑HVCX神经元对传入信息能够进行更迅速地整合.HVCX神经元发放频率通过统计单位时间内动作电位发放个数来表示.在电流钳下给予HVCX神经元500 ms、100 pA刺激,发现右脑HVCX神经元动作电位发放频率较高(图3F).说明雄鸟右脑HVCX神经元的兴奋性强于左脑HVCX神经元.
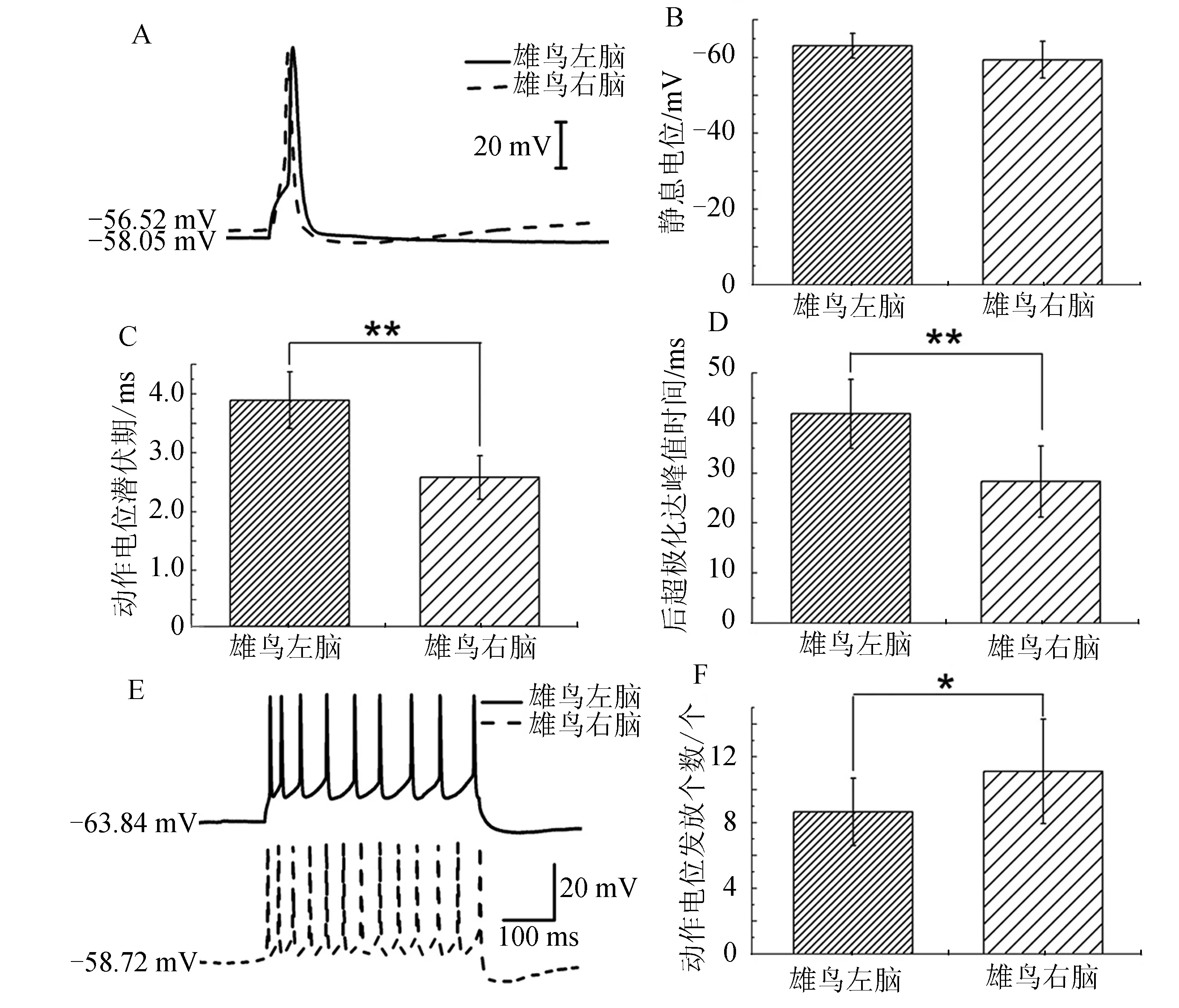
*P<0.05,差异且有显著性;**P<0.01,差异极显著图3 雄鸟左右脑HVCX神经元电生理特性比较Fig.3 The lateral comparison of electrophysiological properties in adult male zebra finches
2.4 雌雄鸟左脑HVCX神经元电生理特性的性别差异
对雌雄鸟左脑HVCX神经元施加5 ms、300 pA电流刺激,将记录到的电生理指标进行统计.结果显示,静息电位(图4B)、动作电位潜伏期(图4C)和后超极化达峰值时间(图4D)均无显著性差异.
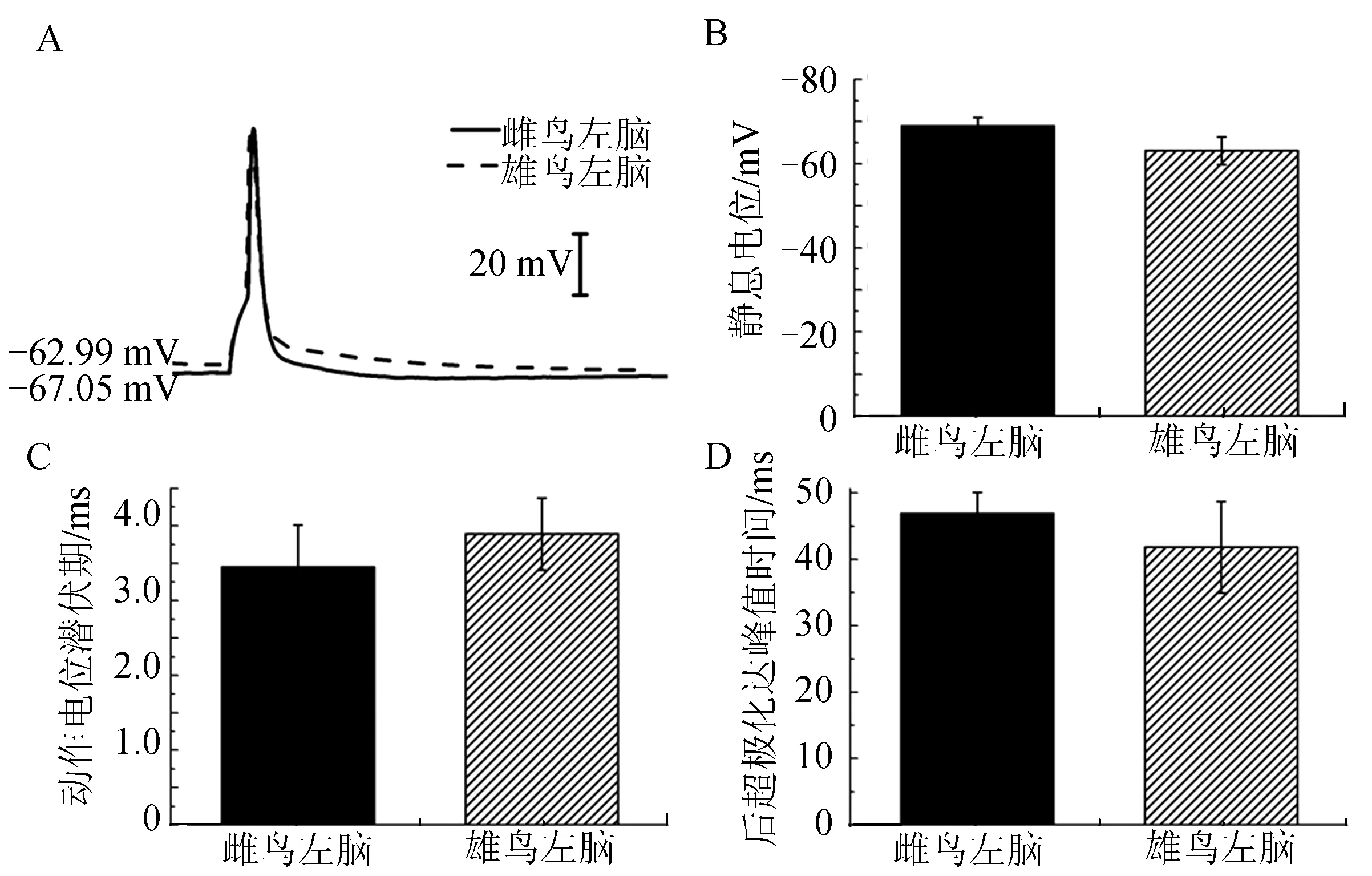
图4 雌雄鸟左脑HVCX神经元电生理特性的性别差异Fig.4 The sexual dimorphism of electrophysiological properties in left brain of adult male and female zebra finches
2.5 雌雄鸟右脑HVCX神经元电生理特性的侧别比较
对雌雄鸟右脑HVCX神经元施加5 ms、300 pA电流刺激,将记录到的电生理指标进行统计.雌雄鸟右脑的静息电位(图5B)无显著性差异.雄鸟右脑的动作电位潜伏期((2.58±0.37) ms,n=5)显著低于雌鸟((3.45±0.56) ms,n=5,P<0.05,图5C).雄鸟右脑的后超极化达峰值时间((27.3±7.1) ms,n=5)低于雌鸟右脑((50.58±3.53) ms,n=5),且达到极显著差异(P<0.01,图5D).在电流钳下给予HVCX神经元500 ms、100 pA刺激,发现雄鸟右脑HVCX神经元动作电位发放频率为((11.1±3.18)个,n=10)显著高于雌鸟右脑((8.71±1.28)个,n=14,P<0.05,图5F).
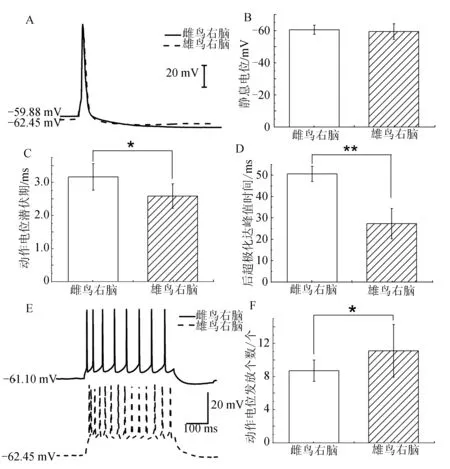
图5 雌雄鸟右脑HVCX神经元电生理特性的性别差异Fig.5 The sexual dimorphism of electrophysiological properties in right brain of adult male and female zebra finches
3 讨 论
雄激素对鸣曲的产生和学习有着至关重要的作用.去势不仅影响鸣曲的学习,还会影响鸣唱核团的大小.雌鸟埋植雄激素会激发鸣曲学习,增加神经元的数量和增大鸣唱核团的体积.虽然雌鸟具有鸣唱核团,但因其体内缺乏雄激素,故其鸣曲学习受阻,只能鸣叫,而不能鸣唱.雌鸟未进行系统的鸣曲学习,HVC核团并不能发出指导鸣曲的指令,尽管雌鸟左右脑的静息电位有显著性差异,但其他指标并无差异,且动作电位发放个数也无差异,说明左右两侧神经元在接受外界刺激或上游信息输入时做出的反应是一致的.
HVC是鸣禽斑胸草雀发声控制通路中的最高级中枢.损毁HVC核团不仅会造成音节丢失,还会导致鸣曲结构发生改变,且均是损毁右侧HVC造成的影响大于损毁左侧HVC造成的影响[10].损毁左侧HVC未对鸣声造成显著影响,而损毁右侧HVC不仅对长鸣和鸣曲的频域特征和声强特征产生显著影响,还会造成主题曲中音节的丢失和主题曲相似度的下降.分别损毁雄鸟左右侧HVC,鸣曲出现去稳定化和音节丢失,主题曲持续时间丢失和音节平均间隔时间均显著增加.但是,损毁左侧HVC对鸣曲的频域声强特征无显著性影响,说明双侧HVC共同负责鸣曲的时域特征.此外,右侧HVC还负责鸣曲的声强特征[6].雄鸟右脑爆发的动作电位延时要低于左脑.动作电位的爆发与钠离子通道的开放有关,当足够多的钠离子通道打开后,局部电位就会增加,达到阈值进而爆发动作电位.而右脑HVCX神经元的动作电位延时短,说明在相同时间内钠离子通道开放数量更多或者相同数量的钠离子通道开放更为迅速,使得其在接受刺激后更易爆发动作电位.雄鸟右脑HVCX神经元的后超极化达到峰值时间比左脑低.该指标的一个重要作用就是与动作电位的发放频率有关.左脑后超极化达到峰值时间长,限制了其动作电位的发放.说明雄鸟右侧HVCX神经元兴奋性高于左侧,提示左右脑HVCX神经元在鸣曲学习中发挥不同的功能,右侧HVCX神经元的兴奋性可能与声强特征的右侧优势有关.
雌雄斑胸草雀均会长鸣,但是雌鸟长鸣基频较低,谐波平坦,而雄鸟长鸣音节时程比较短且稳定,基频较高,在音节中段及尾段出现向下的频率调制.损毁雄鸟左侧HVC前后的长鸣不存在明显的改变,而损毁右侧HVC后,长鸣已不具备习得性长鸣的特征,与雌性长鸣和亚鸣期的长鸣类似.雌雄鸟左脑HVCX神经元电生理特性并无显著性差异,雄鸟右脑HVCX神经元的兴奋性却高于雄鸟左脑,提示雄鸟右侧HVC可能是习得性长鸣与雌性长鸣和亚鸣期长鸣不同的源头.
脑源性神经营养因子(brain-derived neurotrophic factor, BDNF)是一种具有神经营养作用的蛋白质.虽然雌雄两性个体HVC核团中都有脑源性神经营养因子受体TrkB表达,但BDNF只存在成年雄性HVC核团,雌性HVC中并未检测到该因子的存在[11].雌鸟中埋植睾酮可以增加脑源性神经营养因子含量,BDNF通过提高神经元的存活率增加HVC核团神经元数量和核团体积.雌雄鸟性别差异与体内睾酮含量有密不可分的联系,雌鸟埋植睾酮不仅可提高神经元的存活率,而且还会增加神经元胞体之间的间隙连接,提示雄鸟大脑中拥有更复杂的神经网络,雌雄鸟的电生理特性的差异可能与此有关.提示雄鸟右侧HVCX神经元比雌鸟具有更高的兴奋性,利于信息的接收及其向下游核团的传递,有助于鸣曲的学习.
本实验对雌雄两性的侧别差异和性别差异进行了研究.雄鸟个体右侧大脑HVCX神经元兴奋性强于左脑,即存在右侧优势,提示双侧HVC核团除具备共同的鸣曲学习功能外,右侧HVC在鸣曲学习中还发挥其他的作用.雌雄右脑存在性别差异且雄鸟HVCX神经元兴奋性强于雌鸟,雄鸟HVCX神经元能更精准地接收听觉信息以及能将听觉信息有效地进一步传输给下游核团X区,增强AFP通路信息传递能力,从而指导鸣曲的学习.
猜你喜欢
江西农业大学学报(2022年6期)2023-01-31
中国医学影像技术(2022年2期)2022-03-01
幼儿100(2020年29期)2020-10-21
海南师范大学学报(自然科学版)(2020年1期)2020-04-18
旅游纵览(2019年2期)2019-01-31
大自然探索(2017年11期)2017-11-28
小猕猴智力画刊(2016年6期)2016-05-14
唐山文学(2015年1期)2015-11-17
中国实用神经疾病杂志(2012年14期)2012-12-17
