
水稻复合农艺性状QTL剖析
2020-03-25 13:47刘冠明林增顺徐庆国
湖南农业大学学报(自然科学版) 2020年1期
刘冠明,林增顺,徐庆国
水稻复合农艺性状QTL剖析
刘冠明,林增顺,徐庆国*
(湖南农业大学农学院,湖南 长沙 410128)
为了解水稻复合性状的数量性状基因座(QTL),在利用单片段代换系进行QTL鉴定的基础上,剖析了水稻株高QTL与主茎高和穗长QTL,主茎高与倒一节间长、倒二节间长、倒三节间长和倒四及以下节间长QTL,谷粒长宽比QTL与粒长和粒宽QTL,每穗粒数QTL与一次枝梗数和二次枝梗数QTL的关系。结果表明:鉴定出株高QTL的6个单片段代换系中有4个只检测出了主茎高QTL,其加性效应百分率为86.00%~99.55%,有1个只检测出了穗长QTL,其加性效应百分率为48.31%,有1个同时检测出了主茎高QTL和穗长QTL,其中主茎高QTL的加性效应百分率为81.72%,穗长QTL加性效应百分率为18.28%;在检测出主茎高QTL的7个单片段代换系中,有1个只检测出倒一节间长QTL,有2个只检测出倒二节间长QTL,有2个检测出倒一节间长QTL和倒二节间长QTL,有2个只检测出倒三节间长QTL;不同的单片段代换系中检测出的节间长的QTL加性效应百分率变化范围为-128.62%~172.07%;7个检测出谷粒长宽比QTL的单片段代换系中,有5个只检测出粒长QTL,1个只检测出粒宽QTL,1个同时检测出了粒长QTL和粒宽QTL;检测出每穗粒数QTL的3个单片段代换系中,有2个只检测出二次枝梗数的QTL,有1个同时检测出一次枝梗数QTL和二次枝梗数的QTL。这些结果表明,代换片段中如能检出复合性状QTL,也可以检出其构成性状QTL;复合性状QTL的加性效应的大部分可由其构成性状QTL的综合效应来解析,但相同的复合性状,不同代换片段检出的构成性状QTL不同。
水稻;数量性状基因座;复合农艺性状;构成性状;单片段代换系

随着分子标记的发展和检测方法的成熟,越来越多的数量性状基因座(quantitative traits locus,QTL)被检测出[1–3]。一些性状在数量上存在相关关系或构成关系,不同性状的QTL之间也存在一定的关系;因此,了解QTL之间的关系有利于准确了解QTL的功能,更好地利用QTL。目前的QTL分析,多数限于分析复合性状,如产量、株高、每穗总粒数、粒型等[1–5],或将构成性状当作独立的性状进行QTL分析[6~8],而对复合性状QTL与构成性状QTL之间的关系少有报道。
染色体单片段代换系是一类与受体亲本只存在特定染色体片段差异的材料[9]。利用单片段代换系进行水稻QTL鉴定具有简单、准确的特点,已经在许多作物的QTL鉴定中得到了应用[10–13]。本研究中,在利用单片段代换系进行QTL鉴定的基础上,剖析了水稻株高QTL与主茎高和穗长QTL,主茎高QTL与倒一节间长、倒二节间长、倒三节间长、倒四及以下节间长QTL,谷粒长宽比QTL与粒长、粒宽QTL,每穗粒数QTL与一次枝梗数和二次枝梗数QTL的关系,旨为研究水稻复合性状与构成性状间的关系提供参考。
1 材料与方法
1.1 材料
12个单片段代换系,其中,6个来源于低脚乌尖,6个来源于窄叶青(表1),分布于7条染色体。受体为台中65。
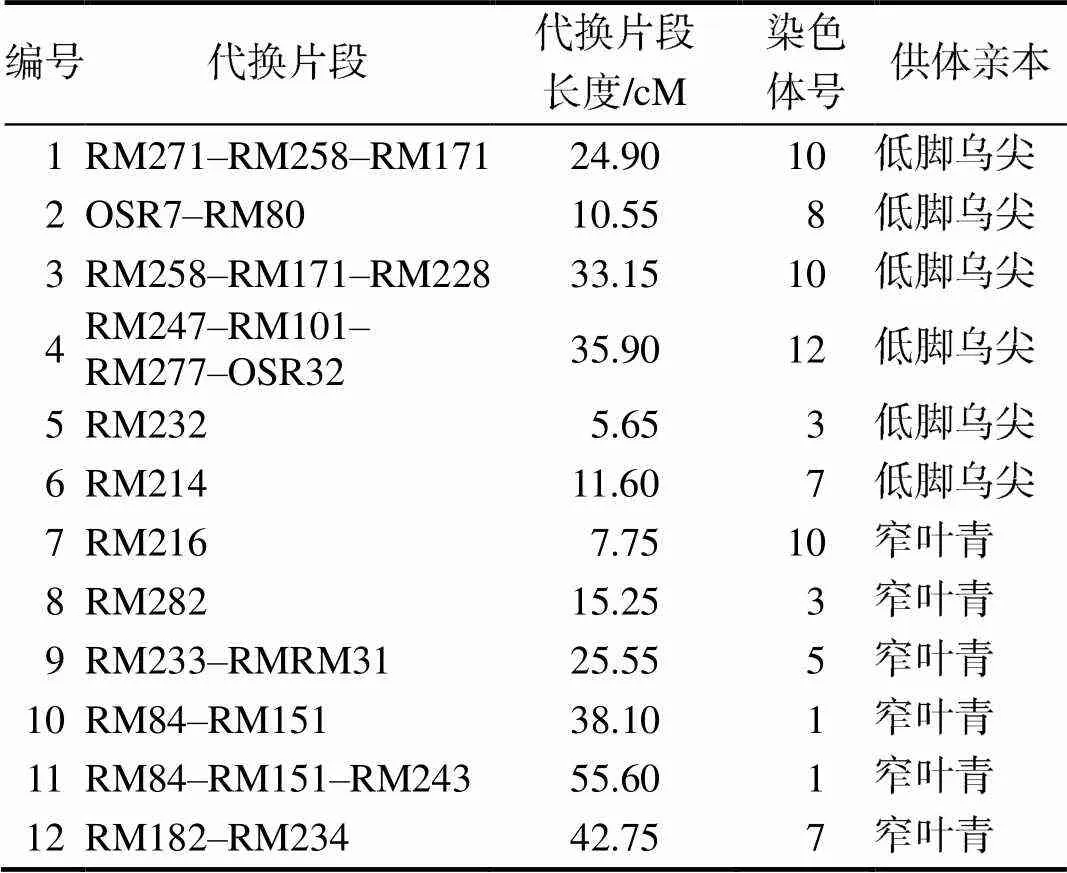
表1 鉴定QTL的单片段代换系
1.2 方法
性状的调查按照文献[14]中的方法进行。QTL的判定参照刘冠明等[9]的方法,通过检验分析单片段代换系与受体亲本台中65(对照)间的差异显著性,以=0.001为标准,判定QTL是否存在,当≤0.001时,QTL存在。鉴定出的QTL参照ESHED等[15]的方法估算加性效应,对与复合性状存在加减关系的构成性状,计算加性效应百分率,加性效应=(纯合单片段代换系表型值-对照表型值)/2。加性效应百分率=(构成性状QTL加性效应/复合性状QTL加性效应)×100%。
2 结果与分析
2.1 株高QTL的剖析
2.1.1株高与主茎高和穗长QTL的关系
株高由主茎高和穗长构成。主茎高由各节间长构成。株高与主茎高、穗长,主茎高与各节间长之间存在线性相加关系。从表2可以看出,检测出株高QTL的6个代换片段中有4个代换片段只检测出了主茎高QTL,主茎高QTL的加性效应占株高QTL加性效应的86.00%~99.55%,效应方向一致;有1个代换片段中只检测出了穗长QTL,穗长QTL的加性效应占株高QTL加性效应的48.31%;有1个代换片段中同时检测出了主茎高QTL和穗长QTL,其中主茎高QTL的加性效应占株高QTL加性效应的81.72%,穗长QTL加性效应占株高QTL加性效应的18.28%。
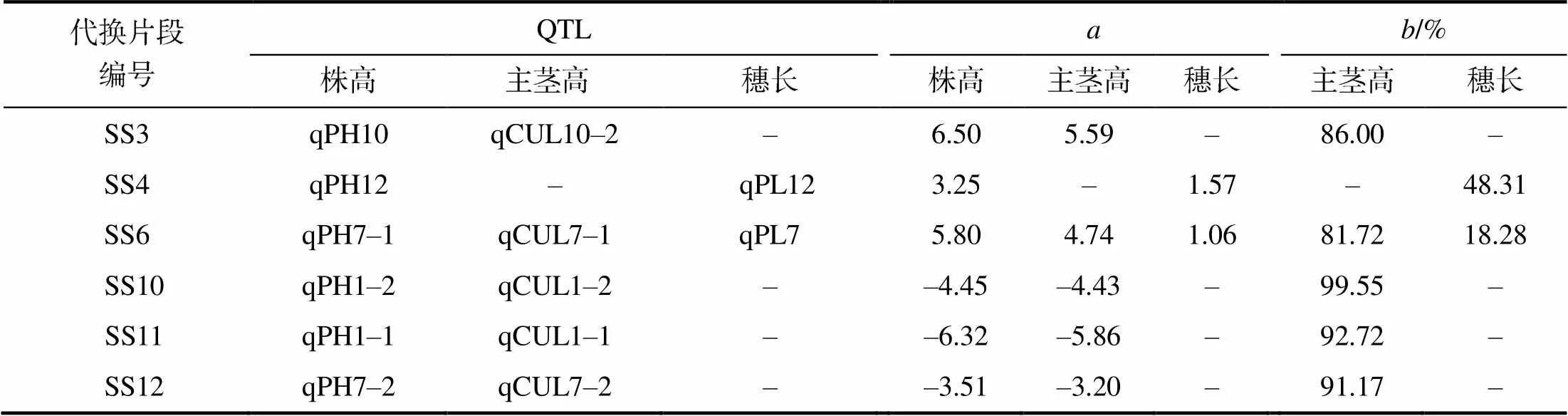
表2 12个代换片段中检出的株高、主茎高和穗长QTL
为相应性状的QTL加性效应;为相应性状的QTL加性效应占株高QTL加性效应的百分率。
2.1.2主茎高与节间长QTL之间的关系
从表3可以看出,在检测出主茎高QTL的7个代换片段中,有1个只检测出倒一节间长QTL,有2个只检测出倒二节间长QTL,有2个检测出倒一节间长QTL和倒二节间长QTL,有2个只检测出倒三节间长QTL。除qLTMI5外,节间长QTL的加性效应方向均与相应的主茎高QTL相同。不同的代换片段中检测出的相同节间长的QTL加性效应占其主茎高QTL加性效应的百分率有差异,如代换片段SS9和SS12中,倒一节间长QTL加性效应占主茎高QTL加性效应的–128.62%和83.75%,而SS11中,倒一节间长QTL加性效应占主茎高QTL加性效应的36.86%。相同代换片段中检测出的不同节间长QTL加性效应占主茎高QTL加性效应的百分率也有差异,如代换片段SS12中,倒一节间长QTL加性效应占主茎高QTL加性效应的83.75%,倒二节间长QTL加性效应占主茎高QTL加性效应的29.06%。
考虑到主茎高的构成性状除倒一节间长、倒二节间长、倒三节间长外,还有倒四及以下节间长,分析倒四及以下节间长QTL的总加性效应发现,代换片段SS9中倒四及以下节间长QTL的总加性效应方向与倒一节间长QTL的相反,与主茎高QTL的相同,部分解析了主茎高QTL的效应来源。其他代换片段倒四及以下节间长QTL的总加性效应方向与倒一节间长、倒二节间长、倒三节间长QTL方向一致,与主茎高QTL也一致,能解析主茎高QTL部分效应来源(表3)。
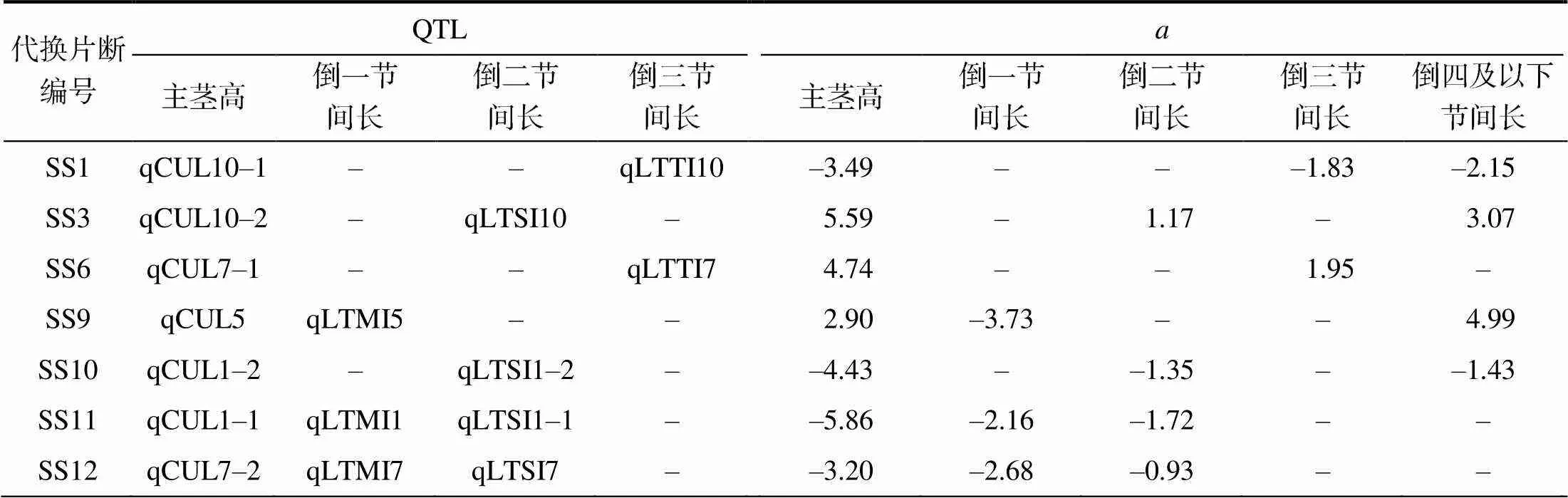
表3 12个代换片段中检出的主茎高与节间长QTL
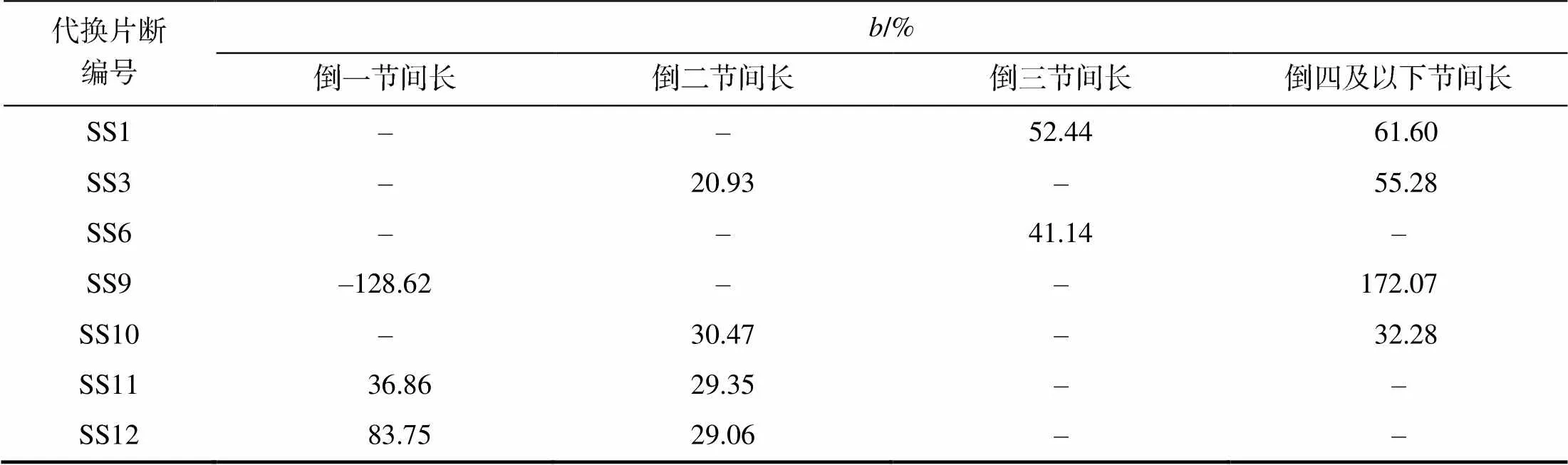
表3(续)
为相应性状的QTL加性效应;为相应性状的QTL加性效应占主茎高QTL加性效应的百分率。
2.2 谷粒长宽比的QTL剖析
在计量上谷粒长宽比与谷粒长成正比,与谷粒宽成反比。代换片段中能检测出谷粒长宽比QTL,也能检测出粒长QTL和粒宽QTL。从表4可以看出,7个检测出谷粒长宽比QTL的单片段代换系中,SS1、SS2、SS4、SS8和SS10中只检测出粒长QTL,SS9只检测出粒宽QTL,SS7同时检测出了粒长QTL和粒宽QTL。谷粒长宽比QTL与粒长或粒宽QTL在效应方向上与计量关系反映的一致。
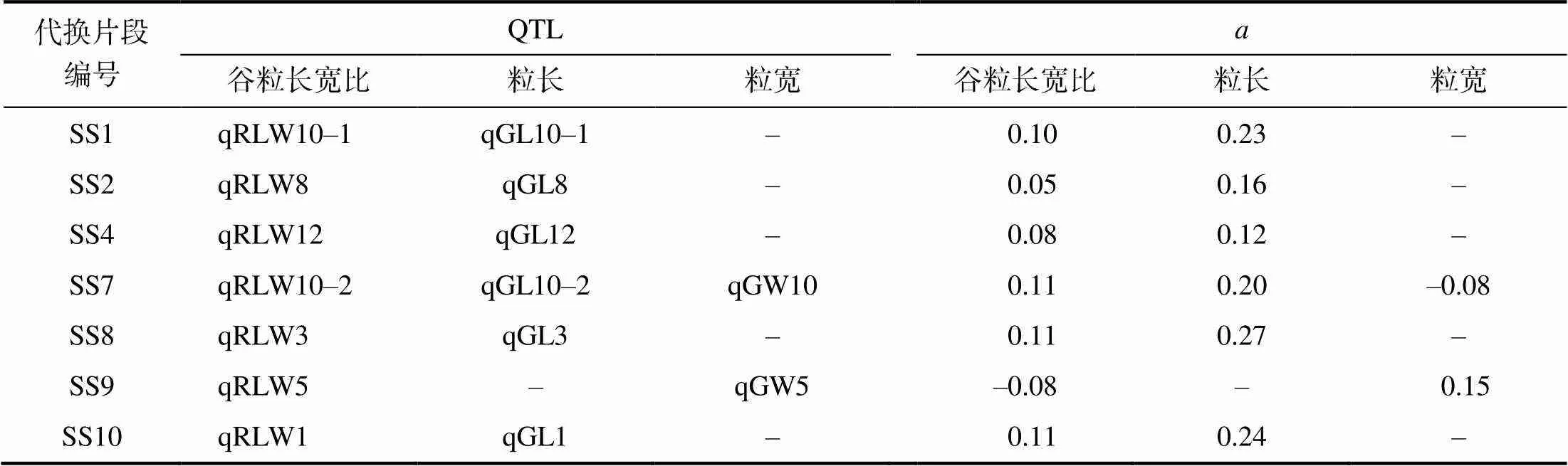
表4 12个代换片段中检出的谷粒长宽比及其构成性状的QTL
为相应性状的QTL加性效应。
2.3 每穗粒数QTL的剖析
一次枝梗数、二次枝梗数可看作是每穗粒数的构成性状,它们之间在计量上成正比关系。
从表5可以看出,代换片段中能检测出每穗粒数QTL,也能检测出一次枝梗数或二次枝梗数的QTL,其中SS3、SS12只检测出二次枝梗数的QTL,SS1中同时检测出一次枝梗数的QTL和二次枝梗数的QTL。每穗粒数QTL与一次枝梗数或二次枝梗数的QTL的效应方向与计量关系反映的一致。
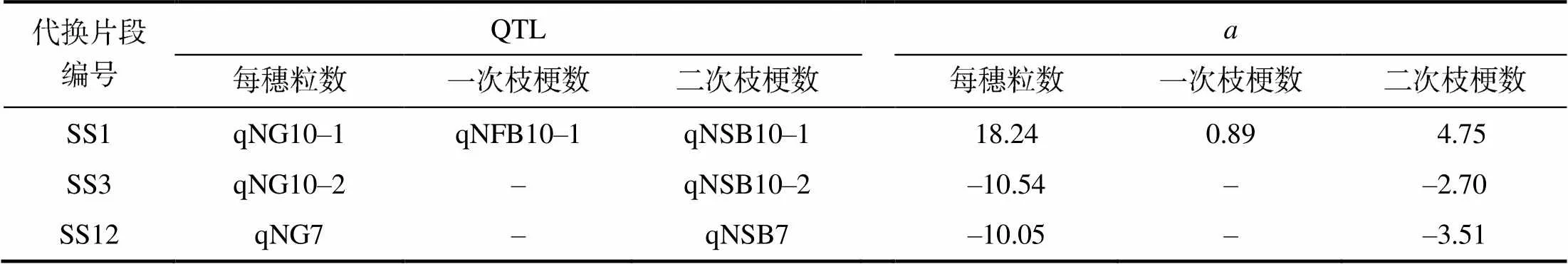
表5 12 个代换片段中检出的每穗粒数及其构成性状的QTL
为相应性状的QTL加性效应。
3 结论与讨论
林鸿宣等[6]利用分离群体对水稻株高及其构成性状QTL进行了分析,发现它们有成簇分布的现象,提出它们可能是同一类QTL,但没有进一步分析株高与其构成性状QTL的具体关系。本研究结果表明,鉴定出株高QTL的6个单片段代换系中有4个只检测出了主茎高QTL,有1个只检测出了穗长QTL,有1个同时检测出了主茎高QTL和穗长QTL。在检测出主茎高QTL的7个单片段代换系中,有1个只检测出倒一节间长QTL,有2个只检测出倒二节间长QTL,有2个检测出倒一节间长QTL和倒二节间长QTL,有2个只检测出倒三节间长QTL;除qLTMI5外,节间长QTL的加性效应方向均与相应的主茎高QTL相同。7个检测出谷粒长宽比QTL的单片段代换系中,有5个只检测出粒长QTL,1个只检测出粒宽QTL,1个同时检测出了粒长QTL和粒宽QTL。检测出每穗粒数QTL的3个单片段代换系中,2个单片段代换系只检测出二次枝梗数的QTL,1个单片段代换系中同时检测出一次枝梗数QTL和二次枝梗数的QTL。说明单片段代换系中复合性状QTL的大部分效应可以由构成性状QTL的综合效应解析,即检测出的复合性状的QTL是其构成性状QTL的综合反映。
本研究结果表明,相同的复合性状在不同位点(代换片段)上检测出的构成性状QTL不同,如主茎高,在有些代换片段中只检测出倒一、倒二或倒三节间长QTL,有些代换片段中能同时检测出倒一节间长与倒二节间长QTL;检测出的相同构成性状的QTL效应大小也有所不同,有些构成性状QTL的效应方向与复合性状QTL相反,如代换片段SS9中检测出的主茎高QTL与倒一节间长QTL。说明不同位点上检测出的相同复合性状的QTL,是不同构成性状QTL之间综合作用的反映;因此,了解复合性状的构成性状QTL,对准确了解复合性状QTL的作用及改良复合性状具有重要意义。
利用传统的材料进行QTL分析时,一个复合性状可同时在多个区间上检测出QTL,这些QTL之间存在互作[16],即使同时分析复合性状与构成性状的QTL,也难以准确分析检测出的复合性状QTL与构成性状QTL之间的关系[17]。单片段代换系中只含有1个外源片段,在鉴定出QTL的同时,实现了QTL的初定位,能同时分析多个性状,遗传背景干扰小,鉴定和定位结果准确可靠[11]。利用单片段代换系分析复合性状与构成性状QTL时,分析的是同一个区间(代换片段)上的关系,这样的结果更能反映真实情况。从本研究结果看,4种复合性状QTL大部分能由其构成性状QTL解析,不能解析的效应可能是由于QTL鉴定时标准过于严格(=0.001),部分QTL没能检测出造成的。
1个代换片段中检测出的复合性状QTL,可能是1个构成性状QTL的反映,也可能是多个构成性状QTL之间互作结果的反映。同一个复合性状的多个构成性状的QTL之间的关系以及同一定位区间检测出的多个QTL之间的关系需要作进一步分析,可将一个代换片段中的QTL分割成单个孟德尔因子,利用单片段代换系构建含更小代换片段的亚代换系,将一个较大代换片段上的多个QTL分割成单个孟德尔进行分析[18]。
[1] XIAO J H,GRANDILLO S,AHN S N,et al.Genes from wild rice improve yield[J].Nature,1996,384:223–224.
[2] YOON D B,KANG K H,KIM H J,et al.Mapping quantitative trait loci for yield components and morphological traits in an advanced backcross population betweenand thecultivar Hwaseongbyeo[J].Theoretical and Applied Genetics,2006,112(6):1052–1062.
[3] 董骥驰,杨靖,郭涛,等.基于高密度Bin图谱的水稻抽穗期QTL定位[J].作物学报,2018,44(6):938–946. DONG J C,YANG J,GUO T,et al.QTL mapping for heading date in rice using high-density Bin map[J].Acta Agronomica Sinica,2018,44(6):938–946.
[4] LI Z K,PINSON S R M,STANSEL J W,et al. Identification of quantitative trait loci (QTLs) for heading date and plant height in cultivated rice (L.)[J].Theoretical and Applied Genetics,1995,91(2):374–381.
[5] 林鸿宣,闵绍楷,熊振民,等.应用RFLP图谱定位分析籼稻粒形数量性状基因座位[J].中国农业科学,1995,28(4):1–7. LIN H X,MIN S K,XIONG Z M,et al.RFLP mapping of QTLs for grain shape traits in Indica rice(L.subsp.)[J].Sientia Agricultura Sinica,1995,28(4):1–7.
[6] 林鸿宣,庄杰云,钱惠荣,等.水稻株高及其构成因素数量性状基因座位的分子标记定位[J].作物学报,1996,22(3):257–263. LIN H X,ZHUANG J Y,QIAN H R,et al.Mapping QTLs for plant height and its components by molecular markers in rice (L.)[J].Acta Agronomica Sinica,1996,22(3):257–263.
[7] 徐建龙,薛庆中,罗利军,等.水稻粒重及其相关性状的遗传解析[J].中国水稻科学,2002,16(1):6–10. XU J L,XUE Q Z,LUO L J,et al.Genetic dissection of grain weight and its related traits in rice (L.)[J].Chinese Journal of Rice Science,2002,16(1):6–10.
[8] ZHANG Y X,ZHOU L Y,SHEN X H,et al.Genetic dissection of yield traits in super hybrid rice Xieyou9308 using both unconditional and conditional genome-wide association mapping[J].Scientific Reports,2017,7:824.
[9] 刘冠明,李文涛,曾瑞珍,等.水稻亚种间单片段代换系的建立[J].中国水稻科学,2003.17(3):201–204. LIU G M,LI W T,ZENG R Z,et al.Development of single segment substitution lines(SSSLs) of subspecies in rice[J].Journal of Rice Science,2003,17(3):201–204.
[10] 刘冠明,李文涛,曾瑞珍,等.水稻单片段代换系代换片段的QTL鉴定[J].遗传学报,2004,31(12):1395–1400. LIU G M,LI W T,ZENG R Z,et al.Identification of QTLs on substituted segments in single segment substitution lines of rice[J] Acta Genetica Sinica,2004,31(12):1395–1400.
[11] LIU G F,ZHU H T,ZHANG G Q,et al.Dynamic analysis of QTLs on tiller number in rice (L.) with single segment substitution lines[J].Theoretical and Applied Genetics,2012,125(1):143–153.
[12] 周勇,陶亚军,姚锐,等.利用染色体片段代换系定位水稻叶片形态性状QTL[J].作物学报,2017,43(11):1650–1657. ZHOU Y,TAO Y J,YAO R,et al.QTL mapping for leaf morphological traits of rice using chromosome segment substitution lines[J].Acta Agronomica Sinica,2017,43(11):1650–1657.
[13] 邵迪,李秋萍,吴比,等.利用染色体片段代换系定位水稻主效抽穗期QTL[J].湖南农业大学学报(自然科学版),2009,35(4):344–347.SHAO D,LI Q P,WU B,et al Mapping of a major QTL for heading date in rice using chromosome segment substitution lines[J].Journal of Hunan Agricultural University (Natural Sciences) 2009,35(4):344–347.
[14] GB/T 19557.7—2004 植物新品种特异性、一致性和稳定性测试指南:水稻[S].GB/T 19557.7—2004 Guidelines for the conduct of tests for distinctness, uniformity and stability: Rice (L.) [S].
[15] ESHED Y,ZAMIR D.An introgression line population ofin the cultivated tomato enables the identification and fine mapping of yield-associated QTL[J].Genetics,1995,141(3):1147–1162.
[16] LUO L J,LI Z K,MEI H W,et al.Overdominant epistatic loci are the primary genetic basis of inbreeding depression and heterosis in rice Ⅱ.Grain yield components[J]. Genetics,2001,158:1755–1777.
[17] 胡文明.作物复杂性状QTL定位相关的几个问题的探讨[D].扬州:扬州大学,2014. HU W M.Discussion of several issues related to QTL mapping of complex traits in crops[D].Yangzhou:Yangzhou University,2014.
[18] MONFORTE A J,TANKSLEY S D.Fine mapping of a quantitative trait locus (QTL) fromchromosome 1 affecting fruit characteristics and agronomic traits:breaking linkage among QTLs affecting different traits and dissection of heterosis for yield[J].Theoretical and Applied Genetics,2000,100(3/4):471–479.
Dissection of QTLs for agronomic complex traits in rice
LIU Guanming,LIN Zengshun,XU Qingguo*
(College of Agronomy, Hunan Agricultural University, Changsha, Hunan 410128, China)
In order to understand quantitative traits loci (QTLs) for complex traits in rice, in this study, the relationships, between the QTL of plant height and the QTL of main stalk height and panicle length, between the QTLs main stem height and updown 1, updown 2, updown 3, updown 4 and lower internode length, between the QTLs of grain length to width ratio and the QTLs of grain length and grain width, and between the QTLs of grain number per panicle and the QTL of primary branch number and secondary branch number. The results showed that: (1) 4 of 6 SSSLs detected were QTLs of main stem height with the percentage of additive effect, 86.00%-99.55%. One of 6 was QTL for panicle lengt with the percentage of additive effect, 48.31%. And, one was the QTLs for both main stem height and panicle length, with the percentage of additive effects, 81.72% and 18.28% respectively. (2) Out of the 7 SSSLs that detected with QTLs for the main stem height, one is with the QTL for updown1 internode length, two with SSSLs QTL for updown 2 internode length, and two with the QTL for updown 3 internode length, Two SSSLs were detected with QTL for updown 1 internode length and updown 2 internode length with the percentage of additive effect of QTLs for internode length ranging from –128.62% to 172.07% in different SSSLs. (3) Out of the 7 SSSLs that detected QTLs for grain length-to-width ratio, 5 SSSLs were the QTLs for grain length. (4) In three SSSLs detected as QTLs for grain number per panicle, 2 were QTLs for the number of secondary branches. In summary, the QTLs for constituent traits can be detected in the condition that QTLs for compound traits can be detected in the substitution segments. And, most of the additive effects of QTLs for compound traits can be analyzed by the combined effects of QTLs for constituent traits, with different QTLs of the constituent trait detected in different substitution segments.
rice; quantitative traits locus; agronomic complex traits; constituent traits; single segment substitution lines
10.13331/j.cnki.jhau.2020.01.001
S511.01
A
1007-1032(2020)01-0001-06
2018–09–25
2019–01–16
农业部超级稻品种选育与推广示范项目(农财发(2011)45号);广东省科技计划项目(2012B020317002)
刘冠明(1971—),男,广东韶关人,博士研究生,主要从事水稻遗传育种研究,gzzklgm@126.com;
,徐庆国,教授,主要从事水稻遗传育种研究,1075324207@qq.com
刘冠明,林增顺,徐庆国.水稻复合农艺性状QTL剖析[J].湖南农业大学学报(自然科学版),2020,46(1):1–6.
LIU G M, LIN Z S, XU Q G. Dissection of QTLs for agronomic complex traits in rice[J].Journal of Hunan Agricultural University(Natural Sciences), 2020, 46(1): 1–6.
http://xb.hunau.edu.cn
责任编辑:毛友纯
英文编辑:柳正
猜你喜欢
中国农学通报(2022年4期)2022-03-02
作物学报(2021年10期)2021-08-05
北方水稻(2021年2期)2021-04-26
作文与考试·高中版(2021年3期)2021-01-14
江苏科技报·E教中国(2020年10期)2020-12-14
江苏科技报·E教中国(2020年6期)2020-07-30
教学月刊·小学数学(2020年4期)2020-05-13
电子技术与软件工程(2018年11期)2018-02-25
湖北农业科学(2017年6期)2017-04-26
小学教学参考(数学)(2017年1期)2017-03-01
