
横断山高山冰缘带两种伞形科特有植物的核型分析*
2020-04-09 08:08孙文远孙文光李志敏
云南师范大学学报(自然科学版) 2020年2期
孙文远, 孙文光, 李志敏
(1.云南师范大学 生命科学学院,云南 昆明 650500;2.云南师范大学 生物能源持续开发利用教育部工程研究中心,云南 昆明 650500)
棱子芹属(Pleurospermum)隶属于伞形科(Apiaceae),为伞形科在欧亚温带分布中的大属[1],其中大部分种类都分布在喜马拉雅地区,特别是横断山区;中国西南横断山区是伞形科植物的一个现代分布中心,而棱子芹属是伞形科分布在横断山高山冰缘带的第一大属[2-3].该属在全球共50种,中国有39种,其中22种为我国特有物种[4].美丽棱子芹(P.amabile)和丽江棱子芹(P.foetens)都分布于中国西南横断山区的高山地带中,多生长于海拔3 600-5 100 m的流石滩和高山灌丛中,均为横断山高山冰缘带特有的伞形科植物[5].
目前关于棱子芹属植物核型方面的研究,仅20种有染色体资料报道;而本属在中国横断山区分布有25种,其中高山冰缘带分布有12种,12个物种中有染色体报道的种数为4种[3,6-7].可见棱子芹属在细胞学方面的报道尚存在一些空白,且该属是伞形科中唯一一个在横断山高山冰缘带分布有10种以上的属,对于研究伞形科在这一地区的起源与演化具有特殊意义.因此本文选取两种棱子芹属在该地区的特有物种进行了研究,除了对棱子芹属提供染色体资料外,还为植物系统与进化等方面提供研究资料.
1 材料与方法
1.1 材料
供试的材料均采自云南滇西北高山冰缘带,采集地详细信息及凭证标本号见表1,对应的凭证标本存于中国科学院昆明植物研究所标本馆(KUN).

表1 棱子芹属供试材料的采集地信息及凭证标本号
1.2 方法
挑选成熟饱满的种子,在4 ℃冰箱中低温层积3个月以上,以打破种子休眠.层积后将种子放在垫有2~3层普通定量滤纸的灭菌培养皿中,在15~20 ℃恒温箱中萌发.待幼嫩根尖长至1 cm左右长时,取生长旺盛的根尖用0.003 mol/L的8-羟基喹啉在室温下避光预处理4 h;然后在4 ℃低温下用卡诺氏固定液(无水乙醇∶冰醋酸=3∶1)固定12~24 h;在60 ℃恒温水浴条件下用1 mol/L HCl解离6~8 min;再用卡宝品红染色22 h以上;以上步骤间均用蒸馏水浸泡材料数分钟并清洗5次以上;染色完成后用常规细胞压片法镜检观察.挑选处于细胞分裂中期、形态清晰且染色体分离程度良好的细胞进行显微拍照,并制成永久封片以便保存.
根据李懋学等人的标准,对每个物种统计60个以上细胞的染色体数目,以85%及以上的恒定数目作为该研究材料的染色体数目[8].每个物种选取6个以上细胞形态清晰、染色体分散良好且数目稳定的中期细胞图片,进行染色体核型的综合分析.之后以国际通用的统计分析方法进行核型分析[9]:按照李懋学等提出的核型标准对核型参数进行分析[8];以Levan等的方法划分染色体类型[10];核型不对称性按照Stebbins的标准进行划分[11];核型不对称系数根据Arano核型不对称指数“As.K%”及Paszko不对称指数“AI”的算法分别计算[12-13].细胞间期核和有丝分裂前期染色体的形态划分按Tanaka的标准[14].
2 结果
美丽棱子芹和丽江棱子芹的体细胞分裂间期核、前期核、中期染色体形态特征、核型图及核型模式图见图1,两个物种的染色体参数及所得的核型参数详见表2和表3.
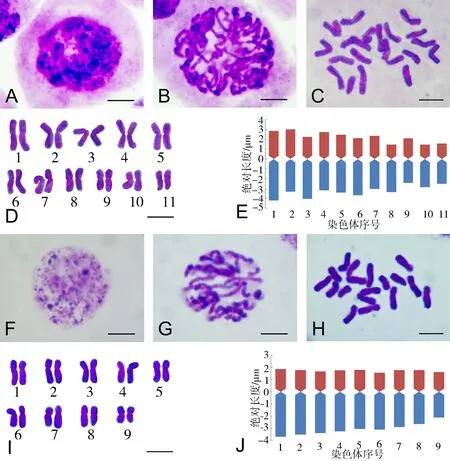
A-E:美丽棱子芹;F-J:丽江棱子芹;E,J:上端为短臂,下端为长臂;标尺=5 μm
图1美丽棱子芹和丽江棱子芹的间期核、前期核、中期染色体、核型图及核型模式图
Fig.1 The resting nucleus,prophase,metaphase chromosomes and haploid idiograms of somatic mitotic in the 2 species ofPleurospermum
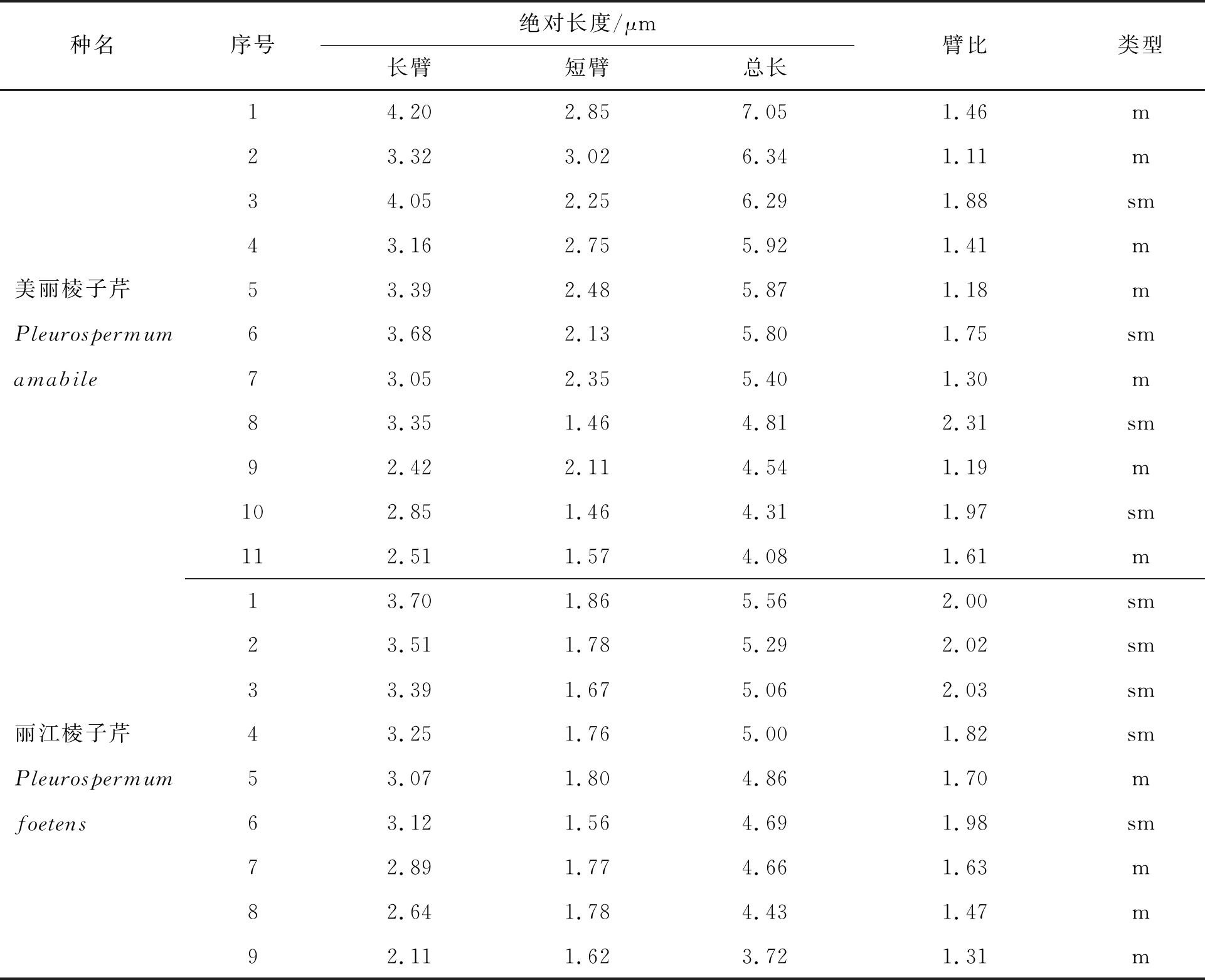
表2 美丽棱子芹和丽江棱子芹的染色体参数

表3 美丽棱子芹和丽江棱子芹的染色体核型参数
CVCI:着丝粒指数变异系数;CVCL:染色体长度变异系数;AI:不对称指数;THL:单倍体染色体组长度总和;MCA:平均着丝粒不对称程度;AsK%:Arano核型不对称指数
研究结果显示美丽棱子芹的体细胞分裂间期核着色较深,染色质几乎均匀地分布在细胞核内,没有明显聚合的染色质小体,根据Tanaka的划分标准为弥散型,见图1-A;图1-B中,前期染色体的异染色质片段位于常染色质片段的相邻位置,按Tanaka的标准属于中间型前期核.丽江棱子芹的体细胞有丝分裂间期核着色较浅,并有一些深染的染色质小体,由染色中心向外逐渐转变为弥散状,根据Tanaka的划分标准为简单型,见图1-F;图1-G中,前期核的异染色质节段和常染色质节段呈相间排列,按Tanaka的标准也属于中间型前期核.
两种棱子芹体细胞有丝分裂中期的染色体长度在3.72~7.05 μm之间变化,依据Lima-de-Faria的标准[15]基本都是中等大小的染色体,见图1-C和图1-H.其Stebbins核型不对称性属于2A型.美丽棱子芹的体细胞核型公式为2n=2x=22=14m+8sm,染色体长度变异为4.08~7.05 μm,臂比范围为1.11~2.31,最长与最短的染色体长度比为1.73,Arano核型不对称指数As.K%=59.55.丽江棱子芹的体细胞核型公式为2n=2x=18=8m+10sm,染色体长度范围为3.72~5.56 μm,臂比变化为1.31~2.03,染色体长度比(最长染色体长度/最短染色体长度)为1.49,Arano核型不对称指数As.K%=63.96.
综合两个物种的核型参数,发现美丽棱子芹和丽江棱子芹均为二倍体,染色体数目2n=18/22,染色体基数x=9/11,所有染色体都属于m或sm型.美丽棱子芹的不对称系数(AI)为2.45,高于丽江棱子芹的不对称系数(AI)值1.11.
3 讨 论
美丽棱子芹和丽江棱子芹的染色体基数、核型不对称性类型以及核型公式均为首次报道.两个物种的Stebbins核型不对称性都属于2A型,在染色体进化上是较为原始的类型.但两者的染色体数目存在差异,结合已有的报道,可以初步确定棱子芹属内种间染色体基数存在差异,具有x=9和x=11两种基数.棱子芹属目前已有的染色体数据支持潘泽惠等人提出的观点,即染色体数目可能对伞形科某些属的属内种间分类有一定意义[16-17].
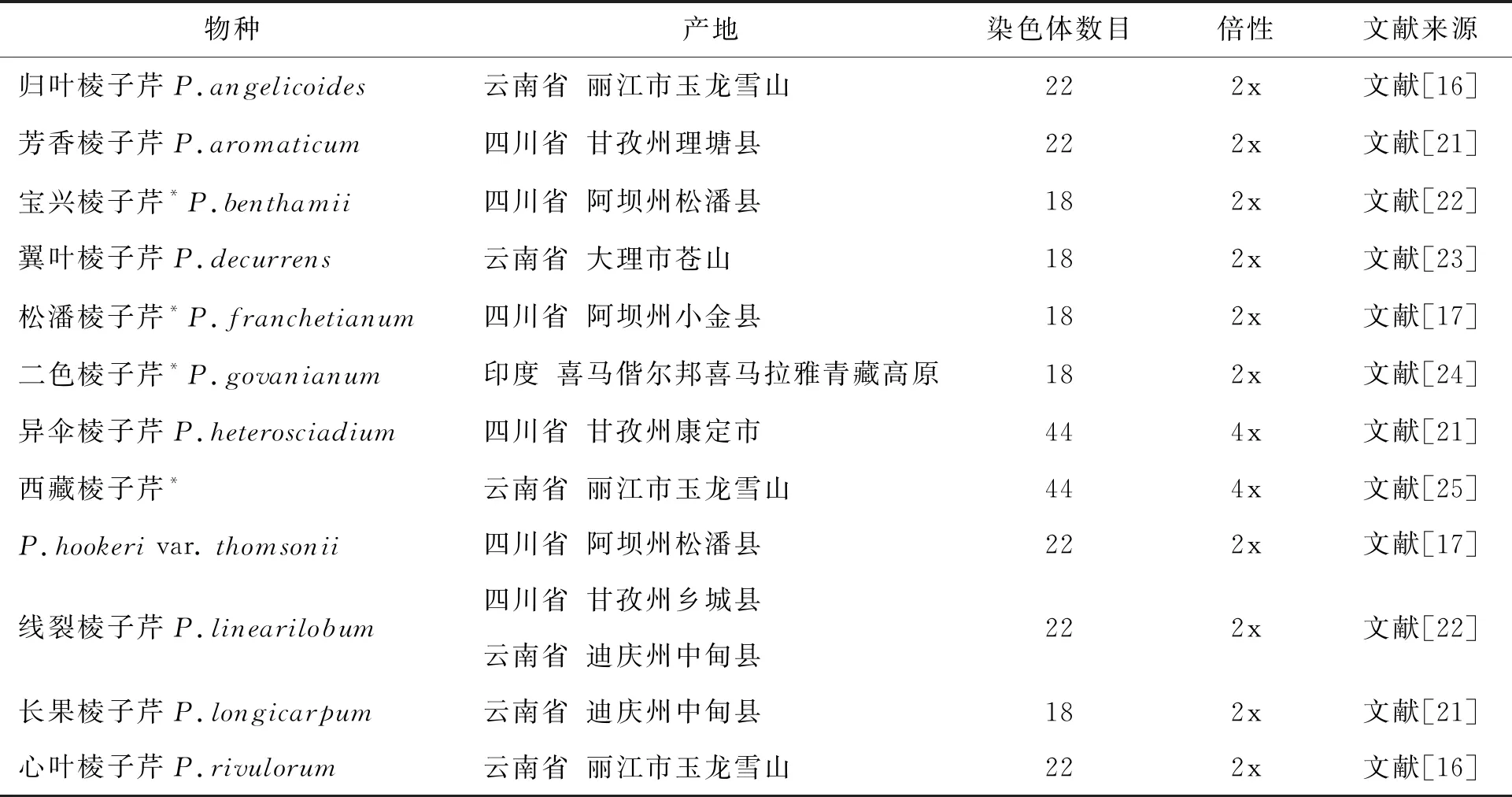
表4 横断山区棱子芹属已有报道的染色体核型和数据来源
*横断山高山冰缘带有分布的物种
前人对伞形科的分子系统学研究均显示棱子芹属不是一个单系类群[18],结合形态学研究[2]及染色体核型数据(表4),对横断山区棱子芹属植物进行染色体基数分析,可以看出其中染色体基数x=9的宝兴棱子芹、翼叶棱子芹、松潘棱子芹、长果棱子芹及本研究中的丽江棱子芹在形态上都具有茎粗壮、伞幅较大、果实大、果棱明显且翅状和总苞片数量多且羽裂等特征;而染色体基数x=11的异伞棱子芹、西藏棱子芹及本研究中的美丽棱子芹在形态上具有茎细弱、伞幅较小、果实小、果棱平直无褶皱或锯齿及总苞片数量少等特征;另外4种染色体基数x=11的归叶棱子芹、芳香棱子芹、线裂棱子芹和心叶棱子芹则是介于上述两组形态特征之间或是具有交叉的形态特征.果实微形态及分子系统学研究显示[18-20],宝兴棱子芹、翼叶棱子芹、松潘棱子芹及丽江棱子芹等10个种属于真正的棱子芹属,而其余的西藏棱子芹、美丽棱子芹、归叶棱子芹、芳香棱子芹、线裂棱子芹和心叶棱子芹等则应划归其他属或是系统位置暂不明确.因此核型数据对于处理棱子芹属下种间分类是具有一定意义的,甚至在处理属间分类上也能提供一定的细胞学参考依据.
对横断山区已有染色体数目报道的棱子芹属植物进行染色体倍性统计发现,该属在横断山区有分布的种仅存在二倍体和四倍体两种倍性,但2n=22,x=11的物种为高基数二倍体,四倍体只出现在异伞棱子芹及西藏棱子芹-丽江玉龙雪山居群中,横断山高山冰缘带分布的12个物种中已有染色体数据的6个种除西藏棱子芹-丽江玉龙雪山居群都是二倍体.从目前的数据来看,这一地区棱子芹属在染色体水平上的演化可能是以二倍体为主的,与这一地区被子植物多倍体比例偏低的研究结论[21]是吻合的.本研究旨在为棱子芹属提供更多的染色体数据和核型特征,也为理解和揭示棱子芹属在横断山区的细胞水平上的分化和适应机制提供理论依据.
猜你喜欢
临床检验杂志(2021年10期)2021-11-24
食品安全导刊(2021年20期)2021-08-30
云南画报(2021年4期)2021-07-22
核农学报(2021年4期)2021-04-15
文苑(2020年12期)2020-04-13
民族音乐(2018年1期)2018-04-18
民族音乐(2017年6期)2017-04-19
———东俄芹属
中国现代中药(2015年9期)2015-09-25
投资与理财(2015年10期)2015-09-10
中国当代医药(2015年9期)2015-03-01
