
基于差示扫描量热技术的板栗胚轴低温保存技术及临界含水量*
2020-04-28 07:09李文清郭素娟解孝满
林业科学 2020年3期
韩 彪 李文清 郭素娟 陆 璐 解孝满
(1. 北京林业大学省部共建森林培育与保护教育部重点实验室 北京 100083; 2. 山东省林木种质资源中心 济南 250102)
顽拗性种子(recalcitrant seed)主要是木本植物种子,一般体积较大,具有较高的经济价值,但顽拗性种子在成熟时多保持较高含水量,对脱水和低温敏感,其保存技术一直是种子保存研究的重要方向(Lietal., 2009; 文彬, 2011; 李磊等; 2016)。顽拗性种子无法长期保存的重要因素之一是脱水敏感性(Waltersetal., 2013; Xiaetal., 2014),由于不同植物、同种植物不同种源种子的脱水耐受性都存在差异,传统研究仅依据萌发的“临界含水量”进行种子顽拗性评价的方法并不完善(Songetal., 2003),出现了同种植物种子的结论迥然不同,甚至不同批次种子的结论也不同,如茶(Camelliasinensis)、小粒咖啡(Coffeaarabica)等都曾被划分为不同种子类型(文彬, 2008)。因此,探索能够客观反映种子水分状态和活力状态的临界含水量,针对不同类型种子研制适宜的保存技术,是顽拗性种子保存的重要研究方向。
板栗(Castaneamollissima)为壳斗科(Fagaceae)栗属(Castanea),是我国重要的木本粮食树种,种植面积和产量均居世界第一(郭素娟等, 2018)。随着板栗产量逐渐增大,其贮藏和保鲜能力不足问题日益凸显, 许多栗农仍采取沙藏法、架藏法等传统方法保存, 一些中小企业虽采用低温贮藏技术,但由于贮前预处理、贮藏温度、相对湿度等参数设置不够科学,贮藏效果也不理想(易善军, 2017)。胚轴是种子萌发的决定因素,是顽拗性种子超纸温保存的核心材料,目前,已有一些关于板栗胚轴超低温保存的报道(郑郁善, 2002; Corredoiraetal., 2004),但保存前临界含水量未有研究,保存后种子萌发率不高。本研究基于差示扫描量热技术(DSC),分析顽拗性种子板栗胚轴的临界含水量,探索板栗种子低温保存技术和胚轴超低温保存技术,以期为板栗等顽拗性种子设施保存提供参考。
1 材料与方法
1.1 试验材料
2018年9月,从山东省济南市西营板栗合作社采集‘烟泉’品种板栗种子,用纱布擦拭干净后自封袋密封包装,2 ℃下保存,备用。
1.2 试验方法
1.2.1 胚轴脱水和含水量测定 选择大小和质量[(10±0.5) g]一致的种子,首先去除种皮,用解剖刀切取含有小块子叶组织的胚轴,将胚轴放在开放的培养皿(直径7 cm)中,然后置于盛有硅胶的干燥器内,使用3个内置风扇增强干燥器内空气循环,快速干燥胚轴。初始含水量测定参考国际种子检验协会(ISTA, 2018)方法,取10个新鲜胚轴称取鲜质量后置于烘箱中(温度103 ℃,烘干17 h)脱水至恒质量,称取干质量后计算含水量=(鲜质量-干质量)/鲜质量×100%,重复3次。在硅胶密闭条件下使胚轴快速失水至50%、40%、30%、25%、20%、15%和10%等梯度含水量,均以新鲜胚轴初始含水量作为对照。
1.2.2 胚轴萌发 参考并改进Corredoira(2004)对欧洲板栗(C.sativa)胚轴的萌发方法,采用恢复培养基和萌发培养基进行板栗胚轴萌发,即将胚轴消毒后,首先接种在恢复培养基上24 h(改良的MS培养基,硝酸盐减半,0.5 mg·L-16-BA,30 g·L-1蔗糖,1 mL·L-1,5 g·L-1琼脂,pH 5.6),然后转接至萌发培养基上(琼脂调整为6 g·L-1,其他成分同恢复培养基)。黑暗条件下培养2周,转移到环境温度为25 ℃的12 h/12 h光周期环境中培养2周,统计萌发率。胚轴萌发20个1组,重复3次。
1.2.3 基于DSC的热力学分析 使用美国TA公司D250系列差示扫描量热仪,对不同含水量胚轴进行热力学测定,以液氮制冷。DSC测定参考Al等(2015)方法,将不同含水量胚轴样品置于铝盘内密封,在微量天平上称质量; DSC分析条件为30 ℃ 保持1 min,以10 ℃·min-1降温至-150 ℃后保持2 min,再以10 ℃·min-1升温至30 ℃。基于DSC软件监测不同含水量胚轴在冷却结晶和加热熔融阶段的热力学数据,分析结晶起始温度、峰值温度和热焓,根据不同含水量胚轴热焓变化计算脱水临界含水量。每个含水量梯度3次重复。
1.2.4 种子低温保存 将采集的板栗种子用纱布擦拭干净后密封保存在自封袋中,置于冰箱保鲜层。根据前期预试验,保存温度设为2 ℃,定期擦拭自封袋内产生的凝结水,每月测定种子发芽率,连续5个月。由于板栗种子存在休眠现象,因此采用组织块萌发方式,即在黑暗条件下培养2周后,转移到环境温度为25 ℃的12 h/12 h光周期环境中培养2周,统计萌发率。种子萌发10个1组,重复3次。
1.2.5 胚轴超低温保存 采用直接冷冻法、程序冷冻法、玻璃化冷冻法和程序玻璃化冷冻法,对不同含水量板栗胚轴进行超低温保存测定。
直接冷冻法: 将胚轴放入冻存管,直接投入液氮中保存。程序冷冻法: 将胚轴放入冻存管,置于-80 ℃冰箱24 h,取出后迅速投入液氮中保存。玻璃化冷冻法: 将胚轴放入加有玻璃化冷冻保护剂PVS2(30%甘油+15%乙二醇+15%二甲亚砜+0.4 mol·L-1蔗糖)的冻存管内1 h,之后迅速投入液氮中保存。程序玻璃化冷冻法: 先将胚轴放入加有玻璃化冷冻保护剂PVS2的冻存管内1 h,然后置于-80 ℃冰箱24 h,最后投入液氮中保存。各种冷冻方法均在液氮中保存24 h。
解冻程序: 取出液氮保存24 h的胚轴,将冻存管置于40 ℃水浴中解冻5 min,含有PVS2的胚轴用无菌水洗涤3次,每次5 min。所有胚轴进行萌发测定,以未经液氮保存的不同含水量胚轴萌发为对照(CK),每组20个胚轴,重复3次。
1.3 数据分析
采用Excel和ANOVA对胚轴萌发和DSC数据进行统计,使用差示扫描量热仪(DSC)TRIOS V4软件(TA,USA)对热力学曲线数据进行分析。
2 结果与分析
2.1 不同含水量胚轴萌发情况
由图1可知,当胚轴含水量从初始的61%降至25%时,胚轴萌发率未明显下降; 当含水量降至20%时,萌发率降至82.1%,与鲜样相比下降18.8%,说明板栗胚轴活力开始下降的临界含水量在20%~25%之间; 当含水量降至10%时,萌发率降至25.6%,大部分胚轴失去萌发能力,与正常种子脱水至含水量10%以下不影响萌发率的现象成鲜明对比。
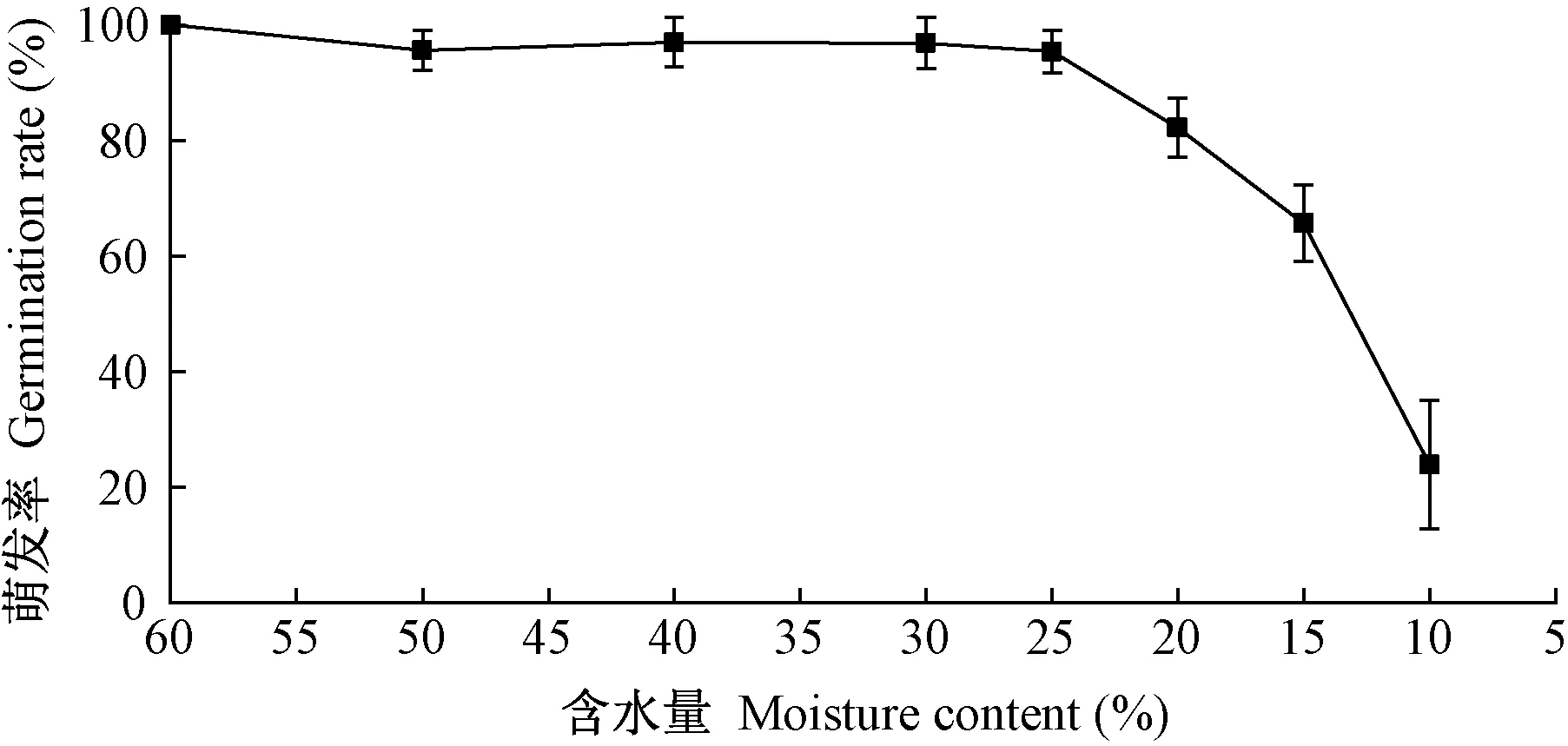
图1 不同含水量胚轴的萌发率Fig.1 Germination rate of embryo axis at different moisture content
2.2 热力学特征分析
DSC热分析显示,随着胚轴含水量降低,结晶起始温度、峰值温度和热焓均呈规律性下降趋势,二者呈显著正相关 (相关系数分别为r1=0.86、r2=0.83、r3=0.99)(图2、表1)。此外,冷却结晶阶段峰值温度低于加热熔融阶段,说明不同含水量胚轴降温过程中形成结晶的温度低于升温过程中结晶融解的温度,胚轴在放热和吸热过程中存在结晶温度差异。
由表1可知,在冷却结晶阶段,未经脱水的胚轴结晶起始温度为-10.6 ℃,峰值温度为-12.0 ℃,热焓为170.2 J·g-1dw,对应的胚轴含水量为61%。当含水量降至25%时,结晶起始温度降至-37.2 ℃,峰值温度降为-44.1 ℃,热焓仅为5.0 J·g-1dw,这说明在脱水过程中可结晶水含量急剧下降,结晶温度显著降低,因此,胚轴含水量在一定范围内越低时产生的结晶伤害风险越小,越有利于种子的低温保存。 当胚轴含水量降至20%及以下时,没有结晶峰产生。加热熔融阶段热力学特征也符合这一规律。
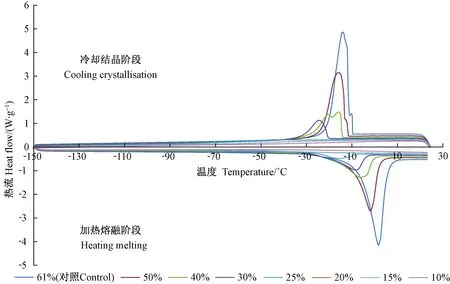
图2 不同含水量胚轴热分析Fig.2 Thermograms of embryo axes at different moisture content
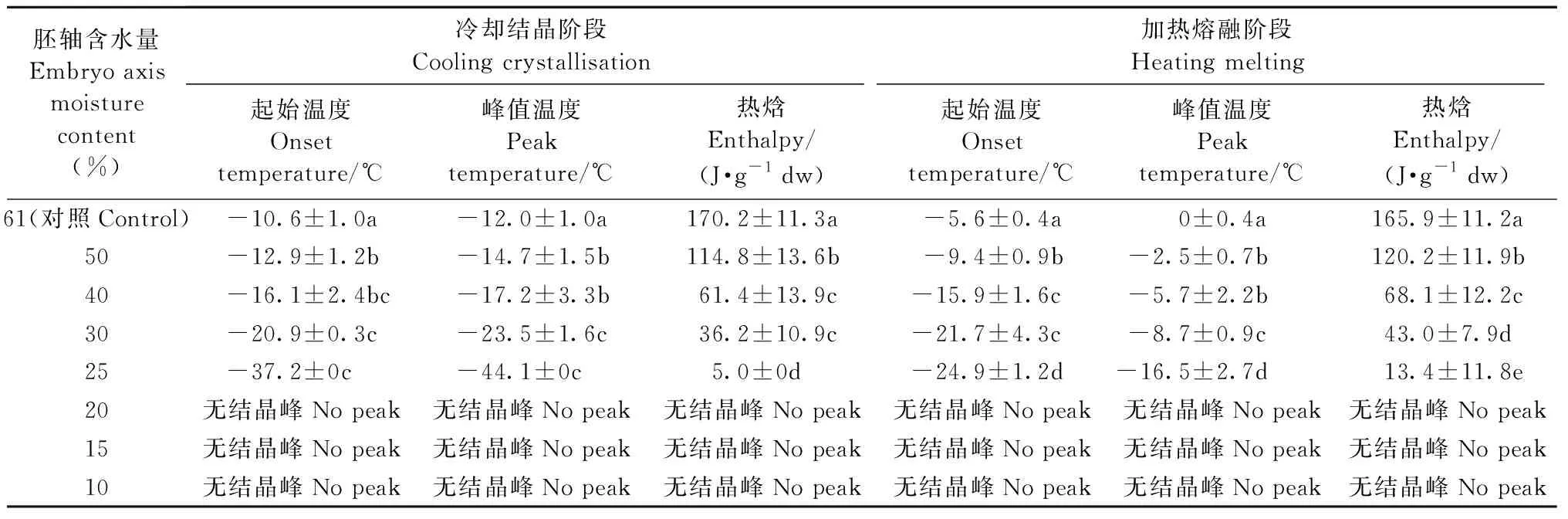
胚轴含水量Embryo axis moisture content(%)冷却结晶阶段Cooling crystallisation加热熔融阶段Heating melting起始温度Onset temperature/℃峰值温度Peak temperature/℃热焓Enthalpy/(J·g-1 dw)起始温度Onset temperature/℃峰值温度Peak temperature/℃热焓Enthalpy/(J·g-1 dw)61(对照Control)-10.6±1.0a-12.0±1.0a170.2±11.3a-5.6±0.4a0±0.4a165.9±11.2a50-12.9±1.2b-14.7±1.5b114.8±13.6b-9.4±0.9b-2.5±0.7b120.2±11.9b40-16.1±2.4bc-17.2±3.3b61.4±13.9c-15.9±1.6c-5.7±2.2b68.1±12.2c30-20.9±0.3c-23.5±1.6c36.2±10.9c-21.7±4.3c-8.7±0.9c43.0±7.9d25-37.2±0c-44.1±0c5.0±0d-24.9±1.2d-16.5±2.7d13.4±11.8e20无结晶峰No peak无结晶峰No peak无结晶峰No peak无结晶峰No peak无结晶峰No peak无结晶峰No peak15无结晶峰No peak无结晶峰No peak无结晶峰No peak无结晶峰No peak无结晶峰No peak无结晶峰No peak10无结晶峰No peak无结晶峰No peak无结晶峰No peak无结晶峰No peak无结晶峰No peak无结晶峰No peak

图3 不同含水量胚轴冷却结晶(A)和加热熔融(B)阶段的热焓Fig.3 The enthalpy of cooling crystallisation (A) and heating melting (B) of embryonic axes at different moisture content
2.3 临界含水量
种子(胚)低温保存的主要伤害来自于细胞内外自由水结晶(冰晶)对细胞膜的破坏(Hamiltonetal., 2009; Aletal., 2015),因此在低温条件下保存板栗胚轴时应尽量避免产生结晶伤害。本研究以冷却结晶(图3A)和加热熔融(图3B)阶段的胚轴含水量为横坐标,以对应的热焓为纵坐标,建立热焓图,其中趋势线与X轴交叉点即为结晶临界含水量。在冷却结晶阶段临界含水量为23.9%,在加热熔融阶段临界含水量为21.6%,二者非常接近,且均处于含水量20%~25%区间。由于自由水结晶伤害主要来自冷却结晶阶段,因此推断‘烟泉’品种板栗的临界含水量为23.9%。
2.4 种子低温保存
根据DSC数据推测,将新鲜种子保存在产生冷却结晶温度之上的低温环境中,不会影响种子活力。如图4所示,在2 ℃低温环境下,保存6个月的板栗种子萌发率达93.3%。种子萌发率、胚根和胚芽长度没有显著差异(P>0.9)。在此温度下将板栗种子保存至第2年春天进行播种繁育,可有效解决顽拗性种子板栗的短期保存问题。
2.5 胚轴超低温保存
根据不同含水量胚轴的萌发结果和DSC热力学数据,对不同含水量胚轴进行液氮超低温保存,其萌发率见表2。对于不同方法超低温保存的胚轴,当含水量在20%及以上时,所有方法均没有萌发,说明此时胚轴在超低温保存过程中受到了严重冰冻伤害。当含水量低于20%时,超低温保存的胚轴部分萌发,在4种超低温保存方法中,直接冷冻法和程序冷冻法的萌发率较低,一般低于10%; 玻璃化冷冻法和程序玻璃化冷冻法的萌发率相对较高,在15%~25%之间,说明采用玻璃化冷冻保护液PVS2处理可提高胚轴超低温保存后的成活率。由于含水量较低时(20%以下)胚轴会遭受脱水伤害,含水量较高时(20%以上)胚轴会遭受冷冻伤害,所以寻找一个合适的平衡点,并结合改进保存技术,是板栗种子和胚轴成功保存的关键。

图4 低温保存板栗种子萌发情况Fig.4 Germination of chestnut seed at low temperature storage
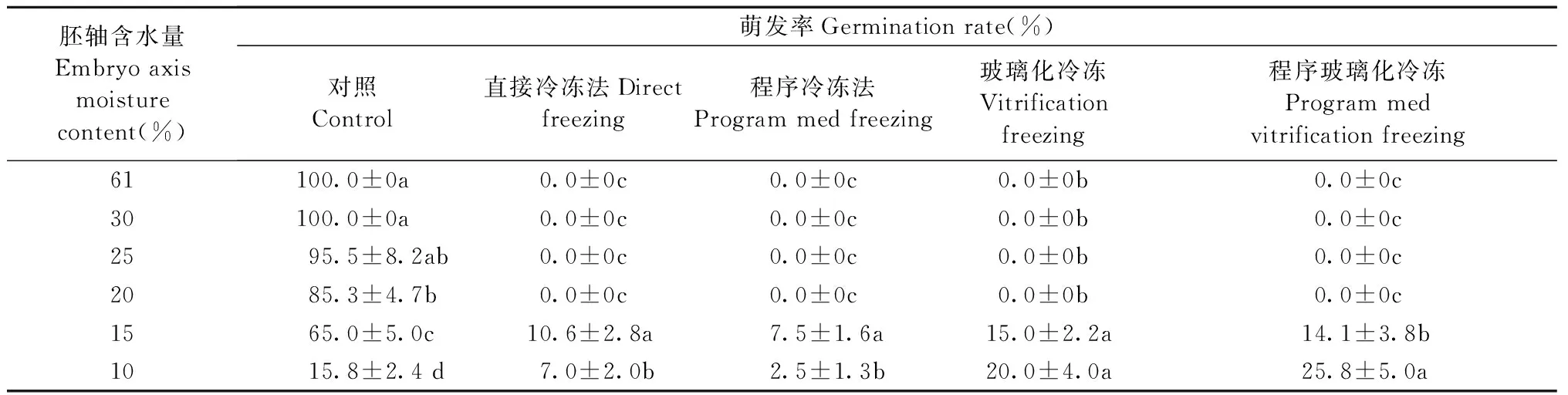
表2 不同含水量胚轴超低温保存的萌发率Tab.2 Germination rate of embryo axes after cryopreservation at different moisture content
3 讨论
顽拗性种子保存难度大,其主要原因是其脱水敏感性,无法像正常性种子那样耐受脱水,种子(胚)含水量是决定顽拗性种子生活力及保存寿命的关键因素 (Xiaetal., 2012; Jinetal., 2018)。壳斗科植物种子是温带地区常见的顽拗性种子,Xia等(2014)比较中国和美国4种栎(Quercus)类顽拗性胚轴耐逆性影响因素,认为胚芽比胚根在切除部位、脱水耐性和低温耐性方面更加敏感。陶月良等(2004)研究板栗种子成熟前后脱水敏感性与可溶性糖、蛋白质之间的关系,发现子叶和胚轴的可溶性蛋白谱带存在较大差异,子叶含有较多的热稳定蛋白。宗梅等(2010)研究不同发育阶段板栗种子的脱水敏感性,结果表明,板栗开花后90天种子中淀粉降解,可溶性糖含量增加,胚轴中可溶性蛋白也大量降解,推断种子已转入萌发状态,由此认为脱水敏感性与细胞膜透性并不完全相关。然而,Chen等(2018)对板栗、豌豆(Pisum)等多种顽拗性种子和正常种子对比分析后发现,细胞膜中磷脂酶Dα1介导的磷脂酸缺失是脱水诱导种子活力丧失的决定因素。可见,目前研究结果还未形成一个成熟的理论体系阐释顽拗性种子脱水敏感性机制,而研究重点又多集中于种子发育阶段和机制研究,对顽拗性种子临界含水量探索较少且观念不一,因此,需在种子萌发基础上,结合DSC等技术手段,优化临界含水量的测定方法。
差示扫描量热技术(DSC)是分析种子和胚轴水分热力学变化的有效工具。种子或胚轴脱水过程中有 结晶起始温度、结晶峰和未冻结含水量3个关键点,结晶峰的形成意味着胚轴组织冰晶伤害(Horetal., 2005; Nadarajanetal., 2008; Aletal., 2015)。本研究基于DSC,结合种子脱水萌发情况,测定‘烟泉’品种板栗胚轴的临界含水量为23.9%,故在没有特殊保护措施情况下,应避免将胚轴脱水至该含水量以下,以免产生脱水伤害。同时,热力学特征表显示,未经处理的板栗胚轴,冷却结晶平均起始温度为-10.6 ℃,理论上保存温度高于该温度就不会形成冰晶伤害,但是由于-10.6 ℃为平均值,不同个体之间冷却结晶起始温度可能存在差异,为了避免对不同个体的冷冻伤害,且方便生产实践应用(普通冰箱冷藏室),本研究将板栗用自封袋密封保存在2 ℃环境中,定期擦拭凝结水,低温保存6个月的种子萌发率和种苗性状基本无变化,种子生活力不受影响,为短期保存板栗种子提供了可能,该方法简单易行,可用于生产中板栗春季播种育苗的种子保存。利用DSC技术可测定顽拗性种子临界含水量,确定种子最适保存温度和保存含水量,该方法准确、快速、简便,可考虑作为顽拗性种子临界含水量的测定方法。
超低温是顽拗性种子保存的可行途径,对于栗属植物超低温保存研究也有一些报道。Pence(1999)对7属18种温带顽拗性种子进行超低温保存,发现栗、山核桃(Carya)、栎这3个属植物可获得完全愈伤组织。郑郁善等(2002)对比不同冷冻处理和解冻处理流程对板栗胚轴生活力的影响,得出各种解冻方式之间差异较小,含水量20%的胚轴萌发率最高为25%。Corredoira等(2004)对欧洲板栗合子胚和体胚进行超低温保存研究,结果发现合子胚含水量20%~24%时成活率较好,达63%。本研究采用4种冷冻方式对板栗胚轴进行超低温保存,显示使用玻璃化冷冻法的胚轴萌发率明显高于直接冷冻法,说明玻璃化冷冻法更适合板栗胚轴保存。另外,无论是欧洲板栗、中国板栗还是中国板栗的不同品种之间,其超低温保存条件都存在一定差异,且超低温保存后恢复培养成苗率均存在较大幅度下降,说明目前的超低温保存方法还不能长期有效保存板栗资源,仍需进一步探索完善。基于DSC的种子低温保存,可作为现阶段板栗等顽拗性种子短期保存和生产应用的有效途径。
4 结论
板栗胚轴含水量降至20%及以下会对胚轴萌发活力产生影响,属于典型顽拗性种子。基于差示扫描量热技术,结合不同含水量胚轴萌发情况,可有效确定板栗等顽拗性种子的临界含水量,‘烟泉’品种板栗胚轴的临界含水量为23.9%。板栗种子在2 ℃低温密封保存条件下可有效保存6个月以上,其萌发率、胚根和胚芽长度无显著变化。加入冷冻保护剂PVS2可提高板栗胚轴超低温保存后的萌发率。
猜你喜欢
特种结构(2021年6期)2021-12-31
特种结构(2021年6期)2021-12-31
上海农业学报(2021年4期)2021-08-28
陶瓷学报(2020年2期)2020-10-27
中国稻米(2019年6期)2019-12-05
制造技术与机床(2019年4期)2019-04-04
江苏农业科学(2018年24期)2018-02-13
上海农业学报(2017年4期)2017-04-10
江西农业学报(2017年12期)2017-02-03
吉林农业(2016年8期)2016-05-14
