
物质的空间分布状态与时间的函数关系
2020-04-28 08:00王文磊
贵阳学院学报(自然科学版) 2020年1期
王文磊
(武汉大学 生命科学学院,湖北 武汉 430072)
本文用加坐标的方法量化细分了无穷小和无穷大,指出只要能在坐标系中打出坐标点,就一定能用连接坐标点的方法得到连续可导的函数图像和关系式,指出物质的所有物理属性变量都与空间坐标变量和时间变量存在着函数对应关系,将物质与空间和时间彻底联系了起来。
1 可变元连续可导函数
分层无穷小:绝对值比任何一个正实数都还要小的非零量是一层无穷小;绝对值比任何一个一层无穷小的绝对值都还要小的非零量是二层无穷小;绝对值比任何一个二层无穷小的绝对值都还要小的非零量是三层无穷小等等,依此无限类推。
分层无穷大:绝对值比任何一个正实数都还要大的量是一层无穷大;绝对值比任何一个一层无穷大的绝对值都还要大的量是二层无穷大;绝对值比任何一个二层无穷大的绝对值都还要大的量是三层无穷大等等,依此无限类推。
同一层的无穷小或无穷大可以像实数轴一样进行细分,它们与原不包含零的实数轴组成分层实数轴。

图1 分层实数轴
如图1,x轴是不包含零的实数轴,y轴的坐标表示层数(负数表示无穷小,正数表示无穷大),x轴上点A,也就是实数1可以表示为N(1,0),点B表示的无穷小量可以表示为N(1,-1),点C表示的无穷大量可以表示为N(1,1)。1除以无穷小量N(1,-1),等于一个无穷大量N(1,1),用除法表示就是:1÷N(1,-1)=N(1,0)÷N(1,-1)=N(1÷1,0+1)=N(1,1)。任何一条实数轴上的点所表示的量,与无穷大N(1,1)相乘后,就“上升”为高一层实数轴上相同横坐标的点所表示的量,与无穷小N(1,-1)相乘后,就“下降”为低一层实数轴上相同横坐标的点所表示的量。可以只保留x轴和y轴,这样纵坐标就不仅仅局限于整数,可以有像N(72,-0.3)、N(2,9.27)这样的无穷小和无穷大(称为二元无穷小、二元无穷大)。
分层三元无穷小:绝对值比任何一个二元无穷小的绝对值都还要小的非零量是一层三元无穷小;绝对值比任何一个一层三元无穷小的绝对值都还要小的非零量是二层三元无穷小;绝对值比任何一个二层三元无穷小的绝对值都还要小的非零量是三层三元无穷小等等,依此无限类推。
分层三元无穷大:绝对值比任何一个二元无穷大的绝对值都还要大的量是一层三元无穷大;绝对值比任何一个一层三元无穷大的绝对值都还要大的量是二层三元无穷大;绝对值比任何一个二层三元无穷大的绝对值都还要大的量是三层三元无穷大等等,依此无限类推。
继续进行分层定义,可以将坐标从二元扩展至三元、四元,乃至无穷,比如N(72,-0.3,7)、N(2,0.3,7,-9),等等。多元坐标实数的基本运算规则为:(1)从左到右,除第一个坐标外的其他坐标都对应相等的,才能加减;(2)坐标个数相等的才能乘除,乘除时第一个坐标乘除,其他坐标加减;(3)第一个坐标不能为零。例如(X≠0,Z≠0,A≠0):
N(X,Y,W)+N(Z,Y,W)=N(X+Z,Y,W)
N(X,Y,W)—N(Z,Y,W)=N(X—Z,Y,W)
N(X,Y,Z)×N(A,W,R)=N(X×A,Y+W,Z+R)
N(X,Y,Z)÷N(A,W,R)=N(X÷A,Y—W,Z—R)

图2 可变元连续可导函数图像例一
如图2,一元函数图像(线段AO)是连续可导的,一元函数图像(线段CD)也是连续可导的,点C无限趋近于点O,点C和点O之间的距离为无穷小,整体函数图像(折线段AOCD)可不可以也是连续可导的呢?在没有分层无穷小等概念前,点O到点C的区域,没有进一步分析的数学工具,因此无法证明点O和点C之间可以存在光滑曲线段相连接。
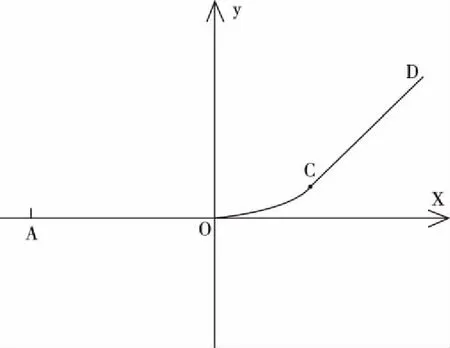
图3 例一的放大图像
有了分层无穷小等概念,可以将点O到点C的距离看成一个无穷小[比如为N(7,-1)],这两点虽然看起来像是重合的,但在这两点之间是可以存在光滑曲线段连接的。如图3,将图像“放大”后[比如两点间距离放大N(1,1)倍],可以看到,可以存在一条光滑曲线段OC将两点连接起来,且曲线段OC与线段AO相切,点O为切点,曲线段OC与线段CD相切,点C为切点。图2的折线段AOCD,可以看成是由线段AO、光滑曲线段OC和线段CD组成,该整体函数图像在点O处和点C处都可导,整体函数图像既处处连续又处处可导,它是连续可导函数。
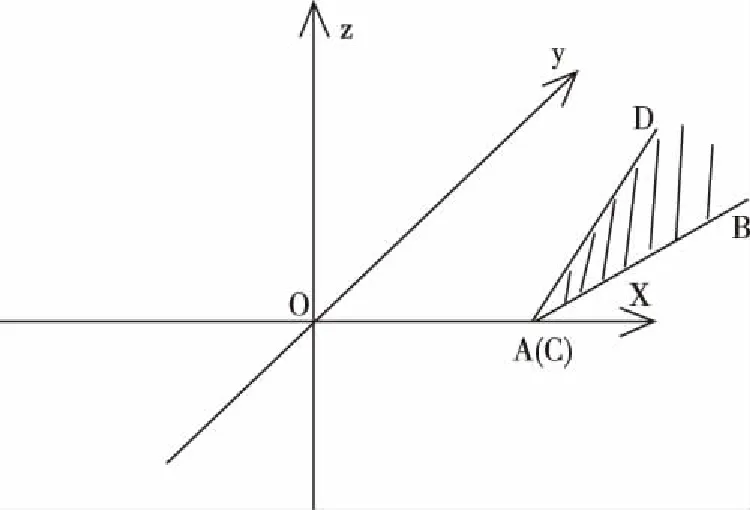
图4 可变元连续可导函数图像例二
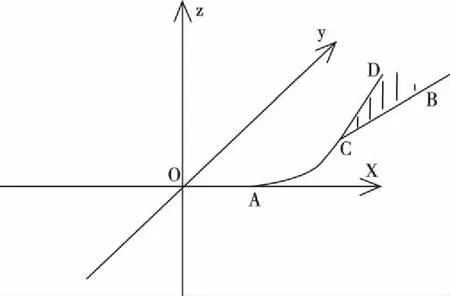
图5 例二的放大图像
如图4,在三维坐标系中一元函数图像(线段OA)是连续可导的,二元函数图像(光滑曲面CBD)也是连续可导的,点C无限趋近于点A。“放大”后如图5,可以存在光滑曲线段AC(一元或二元函数图像),曲线段AC与线段OA相切,点A为切点,曲线段AC在点C的切线,在曲面CBD在点C的切平面上。图4的函数图像,可以看成是由线段OA、光滑曲线段AC和光滑曲面CBD组成,该整体函数图像在点A处和点C处可导,整体函数图像既处处连续又处处可导,它是连续可导函数。
图3表示的函数,其函数变量的个数是固定的(2个),这种函数是固定元函数,图5表示的函数,其函数变量的个数是变化的(2个变为3个),这种函数是变化元函数,固定元函数和变化元函数统称为可变元函数,可变元函数就是指函数变量的个数允许变化的函数。处处连续可导的可变元函数,就是可变元连续可导函数。图3和图5表示的函数就是两个可变元连续可导函数。
2 时间和空间函数原理

时间和空间函数原理:
(1)任何可以测量物理量的事物的运动变化过程,都可以表示为可变元连续可导函数t=G(x1,x2,x3,…,xn),n≥3且n∈Z,Z表示自然数集,t表示时间,x表示事物的各项物理属性,包括但不限于空间三维坐标(x1,x2,x3)。
(2)将组成物质体的微观粒子的质量、电量等物理属性,引入以上函数后可知,任何物质体的运动变化过程,都可以表示为可变元连续可导函数t=G(x1,x2,x3,…,xn),n≥5且n∈Z,x表示微观粒子的各项物理属性,包括但不限于空间三维坐标、质量和电量。
(3)因为空间三维坐标变量加上质量、电量等物理属性的变量,共同确定了物质的空间分布状态,由t=G(x1,x2,x3,…,xn)可知,时间与物质的空间分布状态存在着函数对应关系;或者说,物质不可能脱离空间和时间而单独存在,物质、空间和时间,三位一体紧密联系不可分离。
根据量子力学理论(海森堡测不准原理等),在任一时刻微观粒子的空间坐标,只是表示在这一时刻出现在这里的概率相对最大而已。描述物质体变化规律的可变元连续可导函数,实际是概率函数。
3 生物杂交的函数表达式
生物体的生长变化过程可以用可变元连续可导函数,即生物体函数t=G(x1,x2,x3,…,xn)来表示。生物体在任何时间段的任何组成部分,如卵子、精子、受精卵细胞、胚胎干细胞、某段基因等,都是在“特定时间段”和“特定空间段”的“特定物质组合”,各组成部分所对应的函数图像都是生物体函数图像的局部图像。可变元连续可导函数的图像(例如y=x2的图像),结合分层无穷小等概念可知,该图像(光滑曲线)上任何一部分局部图像(曲线段),都包含了整个函数图像的全部的斜率组合信息(运动变化规律信息)。这就是为什么一个小小的受精卵细胞,包含有生物体全部的生物遗传信息的函数解释。
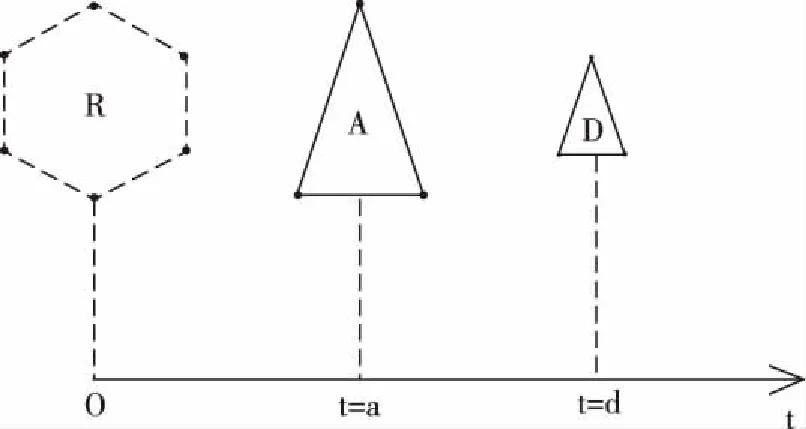
图6 物质杂交结合过程中时间的变化
如图6,将卵子A的诞生时刻和精子D的诞生时刻,看成对应于同一条时间轴的原点O,以A与D杂交结合成功时刻距离原点的时间,为A在杂交结合成功时刻的“生物时间”(t=a),以D与A杂交结合成功时刻距离原点的时间,为D在杂交结合成功时刻的“生物时间”(t=d)。以受精卵细胞R诞生的时刻距离原点的时间,为R的“起点时间”。假设A的生物体函数为t=K(x1,x2,x3,…,xn),A由m个微观粒子组成,A在杂交结合成功时刻的m个坐标点为(a,x1a1,x2a1,x3a1,…,xna1)、(a,x1a2,x2a2,x3a2,…,xna2)、…、(a,x1am,x2am,x3am,…,xnam)。假设D由w个微观粒子组成,D的生物体函数为t=J(x1,x2,x3,…,xn),D在杂交结合成功时刻的w个坐标点为(d,x1d1,x2d1,x3d1,…,xnd1)、(d,x1d2,x2d2,x3d2,…,xnd2)、…、(d,x1dw,x2dw,x3dw,…,xndw)。则有以下m+w个等式:
0=K(x1a1,x2a1,x3a1,…,xna1)-a
0=K(x1a2,x2a2,x3a2,…,xna2)-a
……
0=K(x1am,x2am,x3am,…,xnam)-a
0=J(x1d1,x2d1,x3d1,…,xnd1)-d
0=J(x1d2,x2d2,x3d2,…,xnd2)-d
……
0=J(x1dw,x2dw,x3dw,…,xndw)-d
R对应的生物体函数假设为t=Q(x1,x2,x3,…,xn),已知m+w个坐标点在n维空间坐标系中的坐标点为(x1a1,x2a1,x3a1,…,xna1)、(x1a2,x2a2,x3a2,…,xna2)、…、(x1am,x2am,x3am,…,xnam)和(x1d1,x2d1,x3d1,…,xnd1)、(x1d2,x2d2,x3d2,…,xnd2)、…、(x1dw,x2dw,x3dw,…,xndw),该m+w个n维空间坐标点形成的n维空间图像,在时间轴上的投影数值,就是R诞生时刻对应的“起点时间”。将m+w组变量的数值代入t=Q(x1,x2,x3,…,xn)后,要求存在一个t值,使得:
t=Q(x1a1,x2a1,x3a1,…,xna1)
t=Q(x1a2,x2a2,x3a2,…,xna2)
……
t=Q(x1am,x2am,x3am,…,xnam)
t=Q(x1d1,x2d1,x3d1,…,xnd1)
t=Q(x1d2,x2d2,x3d2,…,xnd2)
……
t=Q(x1dw,x2dw,x3dw,…,xndw)
并且:
0=K(x1a1,x2a1,x3a1,…,xna1)-a
0=K(x1a2,x2a2,x3a2,…,xna2)-a
……
0=K(x1am,x2am,x3am,…,xnam)-a
0=J(x1d1,x2d1,x3d1,…,xnd1)-d
0=J(x1d2,x2d2,x3d2,…,xnd2)-d
……
0=J(x1dw,x2dw,x3dw,…,xndw)-d
可以看出只有t恒等于0时,以上等式才能恒成立。或者说,要使m+w个n维空间坐标系中的坐标点都是某个函数图像上的点,那么这个函数关系式只能为:
0=Q(x1,x2,x3,…,xn)=[K(x1,x2,x3,…,xn)-a][J(x1,x2,x3,…,xn)-d]
所以R的“起点时间”恒定为零,R对应的生物体函数为:
t=[K(x1,x2,x3,…,xn)-a][J(x1,x2,x3,…,xn)-d] ,并且当t=0时,对应的m+w个坐标点(可以称为R的“初始坐标点”)为:(0,x1a1,x2a1,x3a1,…,xna1)、(0,x1a2,x2a2,x3a2,…,xna2)、…、(0,x1am,x2am,x3am,…,xnam)和(0,x1d1,x2d1,x3d1,…,xnd1)、(0,x1d2,x2d2,x3d2,…,xnd2)、…、(0,x1dw,x2dw,x3dw,…,xndw)。 “初始坐标点”表明R的生物体函数关系式,还要受到A和D在杂交结合成功时刻的物质条件的限制。
R的生物体函数t=[K(x1,x2,x3,…,xn)-a][J(x1,x2,x3,…,xn)-d],是其“母亲函数”和“父亲函数”的“结合嵌套”,同时引入了“生物时间”a和d以及“初始坐标点”。这就是在两性繁殖中,“母亲”和“父亲”的生物遗传信息,可以通过自己的后代遗传下去,又会有所变异的函数解释。同一对父母所生的哥哥和弟弟(或姐姐和妹妹),他们的“母亲函数”和“父亲函数”虽然相同,但杂交结合成功时刻卵子的“生物时间”、精子的“生物时间”不相同,“初始坐标点”不相同。因此同一对父母绝对不可能生出一模一样、基因相同的哥哥和弟弟(或姐姐和妹妹)。而同卵双胞胎是由同一个受精卵细胞分裂所得,实际上是同时诞生,“生物时间”相同,“初始坐标点”相同,所以可以一模一样、基因相同。
生物杂交的函数表达式:
(1)假设生物物质A由m个微观粒子构成,A的生物体函数为t=K(x1,x2,x3,…,xn),A在杂交结合成功时刻的“生物时间”为a,在杂交结合成功时刻的m个坐标点为(a,x1a1,x2a1,x3a1,…,xna1)、(a,x1a2,x2a2,x3a2,…,xna2)、…、(a,x1am,x2am,x3am,…,xnam)。
(2)假设生物物质D由w个微观粒子构成,D的生物体函数为t=J(x1,x2,x3,…,xn),D在杂交结合成功时刻的“生物时间”为d,在杂交结合成功时刻的w个坐标点为(d,x1d1,x2d1,x3d1,…,xnd1)、(d,x1d2,x2d2,x3d2,…,xnd2)、…、(d,x1dw,x2dw,x3dw,…,xndw)。
(3)则无论a和d如何取值,A和D经物质杂交结合方式产生的新的生物物质的“起点时间”恒定为零,其生物体函数为:
t=[K(x1,x2,x3,…,xn)-a][J(x1,x2,x3,…,xn)-d]
其m+w个 “初始坐标点”为:
(0,x1a1,x2a1,x3a1,…,xna1)、(0,x1a2,x2a2,x3a2,…,xna2)、…、(0,x1am,x2am,x3am,…,xnam)和(0,x1d1,x2d1,x3d1,…,xnd1)、(0,x1d2,x2d2,x3d2,…,xnd2)、…、(0,x1dw,x2dw,x3dw,…,xndw)。
(4)生物物质杂交结合时,生物遗传信息也会结合并产生变化;杂交结合时有遗传,就一定会有变异,遗传和变异如影随形,不可分离。

图7 细胞的分裂增殖
如图7,点A表示刚受精成功的卵细胞,对应于原点O,表示生命的开始(t=0)。细胞随着时间不断分裂增殖(用分叉表示)的过程,也是第一个受精卵细胞A的组成物质被不断“稀释”的过程。胚胎干细胞是在离O比较近的时间点,从分裂增殖的受精卵细胞中取出的细胞(如图7点B)。因为取出时细胞的分裂增殖次数不多,其内部留存的A的组成物质较多,所以沿着时间轴向右单向发展,它可以形成很多分裂增殖(分叉)。但是,对于已经经过了的分裂增殖,它是不能再重复的,比如点B不能回到点A(时间不能倒流)。对于在取出时与B相分离的其他部分(如C、W、F)的后续分裂增殖,B也是无法表现出来的,因为物质上不允许。胚胎干细胞可以诱导分化成多种组织器官,但不可能诱导分化成完整生物体。因为干细胞永远是受精卵细胞分裂增殖后的局部,其不可能拥有第一个受精卵细胞的全部组成物质。能够分化成完整生物体的,只有受精卵细胞的整体自身。
成年生物体干细胞(如图7点D),因为前面分裂增殖次数多,导致它内部留存的第一个受精卵细胞的组成物质太少,所以沿着时间轴向右单向发展,它后面可以形成的分裂增殖的数量和种类(分化潜力),远远不如胚胎干细胞。干细胞的分化潜力会随着时间的增长(分裂增殖次数增多)而下降。[2]
越早对生物体改造,对未来的影响就越大,因为前面经过的“分叉”较少,可以改变后面更多的“分叉”。对早期基因的修改,比如对受精卵细胞的基因修改,其产生的影响要远远大于,对成年生物体基因修改产生的影响。
iPS细胞(诱导性多功能干细胞),是将受精卵细胞、胚胎干细胞的组成物质(如Oct3/4、Sox2、c-Myc基因片段等),导入“年老”的体细胞中杂交结合。经杂交结合方式形成的iPS细胞的“起点时间”为零(如图7,就是将点D变成点A)。胚胎干细胞的分化多能特性,来自于“年轻”,以及内部含有较多的“第一个受精卵细胞的组成物质”。iPS细胞也具有类似的分化多能特性,是因为其更“年轻”,它的“起点时间”为零,且内部也含有较多的“第一个受精卵细胞的组成物质”(引入的基因片段等)。但iPS细胞所含的“第一个受精卵细胞的组成物质”,不一定比胚胎干细胞多,所以它们之间的分化潜力对比,并无绝对的高下之分。
上海交通大学曾一凡教授等在2009年公布了用iPS细胞培育出活老鼠的实验,认为该实验证明了iPS细胞具有分化“全能性”。该实验过程简单地讲,就是先将“年老”的体细胞(如图7点D),通过物质杂交结合变成iPS细胞(如图7点A,第一次“年龄归零”),然后将与点C、W、F相类似的生物物质(四倍体胚胎),与iPS细胞杂交结合,形成类似受精卵细胞的细胞(第二次“年龄归零”),组合出完整的“第一个受精卵细胞的组成物质”,从时间为零开始继续后面的全部“分叉”,最终发育成完整生物体。它证明的其实是受精卵细胞(或类似受精卵细胞的细胞)分化的“全能性”,而不是iPS细胞分化的“全能性”。
生物克隆本质上与两性繁殖一样,都属于物质的杂交结合,克隆生物的“起点时间”也恒定为零。
实验方案:有人认为生物克隆中,供体的细胞核中的染色体端粒,相对于供体诞生时已经变短了一些,所以供体的细胞核与受体的去细胞核的卵细胞杂交结合后,诞生的克隆生物,其细胞核中的染色体端粒也是短的,因此克隆生物会早衰。但事实并非如此,在杂交结合时会发生复杂的变化,“时间归零”,克隆生物的细胞核中的染色体端粒会变长,恢复到最初的、细胞还没有开始分裂时的长度。也就是说,如果不考虑克隆技术操作造成的生物损伤,克隆生物根本就不会早衰。例如,假设成功杂交结合前的时刻,供体细胞核中的染色体端粒长度为a,克隆生物诞生时刻(即杂交结合成功时刻)细胞核中的染色体端粒长度为b,则通过实验测量可以发现,b>a。
4 基因修改会导致全部生物功能变化的原理
任何一种生物功能对应的函数图像都是生物体函数图像的局部图像。由t=Q(x1,x2,x3,…,xn)=[K(x1,x2,x3,…,xn)-a][J(x1,x2,x3,…,xn)-d]可知,通过物质杂交结合的方式(如各种转基因技术、CRISPR/Cas9技术等)修改基因,会导致变量的对应关系从K(或J)变成Q,所以未来所有的坐标点都会发生变化,会导致所有的局部图像(生物功能)都发生变化。
基因修改会导致全部生物功能变化的原理:
(1)生物物质在任何时间段的任何组成部分,都包含有该生物物质全部的生物遗传信息。
(2)生物物质通过杂交结合的方式产生新的生物物质,物质结合时,遗传信息也会结合并产生变化,所以会产生“牵一发而动全身”的效果,导致全部的生物功能都会产生变化(只是各种变化或大或小、或强或弱罢了)。
(3)通过物质杂交结合的方式对任何基因片段的任何修改,都会使全部的生物功能产生变化(只是各种变化或大或小、或强或弱罢了);仅仅使单一生物功能产生变化的基因片段修改是不存在的。
器官移植与基因修改在本质上一样,都是物质的杂交结合,器官移植后全部的生物功能也都会产生或大或小、或强或弱的变化。临床中移植了器官的人,会产生部分性格和生理的变化,即是与此有关。2018年贺建奎副教授修改人类受精卵细胞基因诞生出婴儿的事件,是希望婴儿对艾滋病病毒有抵抗力。这种愿望是有可能实现的,但问题是:如果出现别的生物功能的改变,导致出现难以预料的灾难怎么办?通过物质杂交结合的方式,人工修改受精卵细胞基因后出生的婴儿,其未来可能的结果只有一种:无论是否具有对艾滋病病毒的抵抗力,其他全部生物功能都会受到或大或小、或强或弱的影响。最终结果是好是坏,答案只能是:事先无法预测。
猜你喜欢
环球人物(2022年4期)2022-02-22
小资CHIC!ELEGANCE(2021年32期)2021-09-18
科学(2020年1期)2020-08-24
当代水产(2019年7期)2019-09-03
中央民族大学学报(自然科学版)(2016年3期)2016-06-27
广西林业科学(2016年2期)2016-03-20
中国教育信息化(2015年10期)2015-08-23
中央民族大学学报(自然科学版)(2015年2期)2015-06-09
小学阅读指南·高年级版(2014年2期)2014-05-27
小说月刊(2014年11期)2014-04-18
