
分子标记辅助选育抗褐飞虱水稻恢复系
2020-05-05 03:14刘百龙张晓丽秦学毅
西南农业学报 2020年3期
冯 锐,郭 辉,陈 灿,刘百龙,张晓丽,秦学毅
(广西农业科学院水稻研究所/广西水稻遗传改良重点开放实验室,广西 南宁 530007)
【研究意义】稻褐飞虱[(NilaparvatalugensStål),简称BPH]是我国及亚洲地区水稻生产中为害最严重的害虫。褐飞虱具有生长周期短、繁殖力强、易迁飞等生活习性,主要通过刺吸危害,造成水稻减产或绝收[1],在我国发生面积达2500万hm2/年,造成巨大经济损失,也成为2003年以来稻米价格上升的主要原因之一[2]。目前,在我国大面积推广的主栽水稻品种中对稻飞虱具有抗性的品种不多,稻田生态系统基本失去了抵御稻飞虱入侵的第一道天然屏障,抗性品种的严重缺乏已成为稻飞虱连续暴发的重要原因之一。分子标记辅助选择(MAS)技术在水稻育种应用中已逐步得到发展和完善,并成为一种重要的育种技术[3-4]。通过分子标记辅助选择有利基因对现有的资源或品种进行遗传改良,已成为抗褐飞虱品种选育的重要途径[5]。抗性育种以抗性基因的鉴定为前提,但是,抗虫鉴定受虫源、环境等影响较大,且耗时费力。利用分子标记辅助选择技术可在早期世代选择抗虫株系,不受虫源和环境影响,可提高育种效率[6-10]。因此,通过分子标记辅助选择结合传统育种技术进行抗褐飞虱水稻恢复系选育,对杂交稻抗性育种的种质资源选择具有重要意义。【前人研究进展】近年来,培育和种植抗虫水稻品种已成为安全、有效实现水稻产业绿色生产和可持续发展、降低农药使用量的途径之一[11],通过分子标记辅助选择,利用有利基因对现有的资源或品种进行遗传改良已经成为行之有效的方法[12]。分子标记选择技术可有效提高目标性状的选择效率,缩短育种周期[13]。在20世纪60年代,Pathak等[14]首次鉴定出栽培稻Mudgo为抗褐飞虱品种。李月鲲等[15]从304份水稻品种(组合)中筛选出抗褐飞虱性能稳定的R373(组合)作为培育抗褐飞虱水稻品种的基础材料使用。张建福等[16]将分子标记辅助选择技术与传统育种结合,创制了米质优、配合力好且抗白背飞虱的新恢复系材料。闫成业等[17]利用分子标记辅助选择改良了水稻恢复系R1005的褐飞虱抗性。罗世友等[18]利用分子标记辅助选育得到抗褐飞虱的改良恢复系。徐鹏等[19]通过分子标记辅助选育,使恢复系R813的稻瘟病、白叶枯病和褐飞虱抗性明显提高。降好宇等[20]利用分子标记辅助选择手段检测携带广谱抗稻瘟病基因Pi9的水稻品系75-1-127,发现其含有褐飞虱抗性基因Bph14和Bph15,可作为抗源应用于水稻褐飞虱抗性育种。李进波等[21]在2006年已利用分子标记辅助选择获得160份目标基因纯合且农艺性状稳定的株系。【本研究切入点】目前,市场上推广的水稻品种中抗褐飞虱品种为数不多,而少有的抗褐飞虱品种中对褐飞虱抗性较差,水稻生产中仍然依赖于化学药剂来防治褐飞虱,但关于以分子标记辅助选择含抗褐飞虱基因水稻品种的研究鲜见报道。【拟解决的关键问题】以高抗褐飞虱水稻品系HS204为母本、恢复系明恢63为父本,通过杂交改良、回交和自交选育,结合分子标记辅助选择和抗虫鉴定,在改良后代中选择褐飞虱抗性强的恢复系材料,为抗褐飞虱杂交稻育种提供良好的种质资源。
1 材料与方法
1.1 试验材料
试验以HS204为供体亲本(母本),其高抗褐飞虱的抗性来源于广西普通野生稻;以水稻恢复系明恢63为受体亲本(父本,轮回亲本);以Ptb33为抗虫对照(CK-R);以TN1为感虫对照(CK-S)。
1.2 试验方法
1.2.1 选育流程 以HS204为母本、恢复系明恢63为轮回亲本进行杂交,经2次回交后多次自交,从BC1F1世代开始进行分子标记辅助选育,在分离群体中筛选含抗褐飞虱基因的植株继续与轮回亲本回交,后代不断自交,选育出抗性基因纯合、农艺性状表现优异的株系,最后进行苗期褐飞虱抗性鉴定。
1.2.2 分子标记 利用所筛选的与抗源HS204抗褐飞虱基因紧密连锁的多态性SSR标记M4-10(F:5′-GTGTAGCTGCTAGGCCGAAC-3′,R:5′-TTCCTTT CGCTACGTTGGAC-3′)和INDEL标记M4-228(F:5′-TGTGCAGCTTTTTCAGGTTG-3′,R:5′-ATGTCGTTCCTTTTGCTGCT-3′),对分离群体中的抗性基因进行跟踪检测,选择携带目标基因的株系。
1.2.3 PCR反应体系和反应程序 参照CTAB法从水稻幼嫩叶片中提取DNA样品。PCR反应体系10.0 μl:DNA模板1.0 μl,10×PCR Buffer(含Mg2+) 1.0 μl,10 mM dNTP 0.2 μl,5 U/μlTaqDNA Polymerase 0.1 μl,10 μM上、下游引物各0.2 μl,ddH2O补足10.0 μl。扩增程序:94 ℃预变性5 min;94 ℃ 40 s,55 ℃ 40 s,72 ℃ 45 s,进行35个循环;72 ℃延伸5 min,4 ℃停止反应。参考李进波等[10]的方法进行电泳和银染检测。
1.2.4 褐飞虱苗期抗性鉴定 供试虫源为广西南宁田间采集混合生物型,在网室用TN1饲养繁殖备用,以褐飞虱生物型II为主,与部分越南九龙江型、孟加拉型和生物型I构成混合生物型。参照苗期集团筛选法将试验材料分为若干组,取各组每份适量种子,催芽后条播于52 cm×33.5 cm×7.5 cm的铝托盘内,每行播20~25 粒,当苗长至3 叶1心时去除小苗、弱苗,保留健壮一致的苗并统计苗数,按每株5~8头接入2~3龄若虫,置于纱网网室中,自然光照,室温25~30 ℃,保持托盘一直有水。在感虫对照品种TN1死亡7 d后调查对照受害症状等级表现,按照表1统计各株系的抗性水平。
1.2.5 改良株系及杂交配组组合主要农艺性状测定 2018年夏季在广西农业科学院水稻研究所试验田种植改良株系与自育不育系1A、2A和10A进行配制的杂交组合,以天优华占为改良株系和杂交组合的对照(CK)。每个组合设3个重复,每小区种植40苗,每行种8株,单株插,密度为16.7 cm×20.0 cm,常规田间管理。记录生育期,在成熟期每小区选取3株,考察株高、单株有效穗、穗长、每穗总粒数、结实率和千粒重等主要农艺性状。
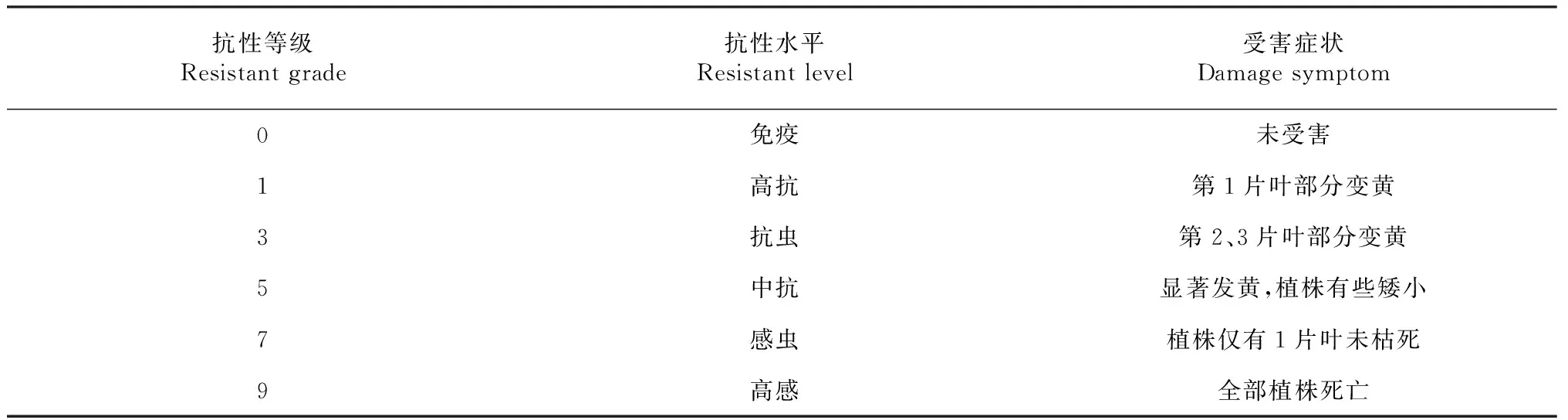
表1 褐飞虱抗性分级标准Table 1 Grade standard of resistance to brown planthopper in Oryza rufipog

图1 MAS改良抗性水稻恢复系的育种路线Fig.1 The technical approach of improving rice resistant restorer line by MAS
1.3 统计分析
试验数据采用Excel 2010进行统计分析。
2 结果与分析
2.1 改良株系的构建及分子标记辅助选择
从图1可看出,杂合单株同时含有母本(供体亲本HS204)、父本(受体亲本明恢63)双亲的2个双条带,阳性纯合单株只有与HS204相同的2个条带,隐性纯合单株只有与明恢63相同的2个单条带。利用分子标记对单株进行检测,共检测到16个抗性基因纯合单株,从中筛选获得RMH-3、RMH-58、RMH-153和RMH-262共4个田间选择农艺性状与受体亲本相似的恢复系单株。

M:200 kb Marker;P1:供体亲本HS204;P2:受体亲本明恢63;F1:杂交一代;泳道1~30:BC2F2代分离群体单株M:200 kb Marker;P1:Donor parent HS204;P2:Recipient parent Minghui 63;F1:First-filial generation;Lane 1-30:Segregation population of BC2F2图2 分子标记检测BC2F2代单株的部分检测结果Fig.2 Detection of molecular marker in partial individuals of BC2F2 population

表2 育成株系对褐飞虱的抗性鉴定结果Table 2 Identification of resistance of restorers against BPH
2.2 改良株系对水稻褐飞虱的抗性鉴定
抗性评价工作在南宁市广西农业科学院温室进行,以感虫品种TN1、抗虫品种PTB33为对照,对选择的4个株系RMH-3、RMH-58、RMH-153和RMH-262进行褐飞虱抗性鉴定。从抗性鉴定结果(表2)可看出,感虫品种TN1全部枯死(9级),4个中选株系RMH-3(3.9级)、RMH-58(1.9级)、RMH-153(3.1级)和RMH-262(1.8级)表现抗和高抗褐飞虱。说明改良株系的抗性较受体亲本明恢63明显提高。
2.3 改良株系及杂交配组的主要农艺性状表现
将选育的4个恢复系改良株系与自育的不育系1A、2A和10A进行配组,获得12个测交组合,对其8个主要农艺性状的分析结果(表3)表明,各测交组合的株高在102.5~120.5 cm,其中1A×RMH-3的株高最矮,明显矮于父本RMH-3,而2A×RMH-262的株高最高,明显高于父本RMH-262;改良株系RMH-3与3个不育系配组的3个组合的有效穗数(分别为7.5、8.5和8.0穗/株)均比父本RMH-3有所增加,改良株系RMH-58与不育系1A、2A配组组合的有效穗数(分别为8.0和7.5穗/株)较父本RMH-58明显增加,而与不育系10A配组组合的有效穗数明显减少;4个改良株系中,除RMH-58外其余3个改良株系的穗长(26.5~27.8 cm)均不同程度长于其父本明恢63,而12个杂交组合中,除2A×RMH-153外其他11个杂交组合的穗长(27.5~32.0 cm)均长于受体亲本明恢63;4个改良株系的每穗实粒数(172.8~233.0粒)均明显少于其父本明恢63,而12个杂交组合的每穗实粒数或多于或少于受体亲本明恢63;4个改良株系的结实率与12个杂交组合无明显差异,而4个改良株系和12个杂交组合的千粒重和单穗重均明显高于受体亲本明恢63;4个改良株系谷粒的长宽比除RMH-58稍小于父母本外其余均介于父母本之间,而大部分杂交组合谷粒的长宽比小于供体亲本HS204,部分大于受体亲本明恢63,部分小于受体亲本明恢63。说明改良株系及其配制的多数杂交组合具有良好的产量表现,部分农艺性状甚至优于受体亲本明恢63。
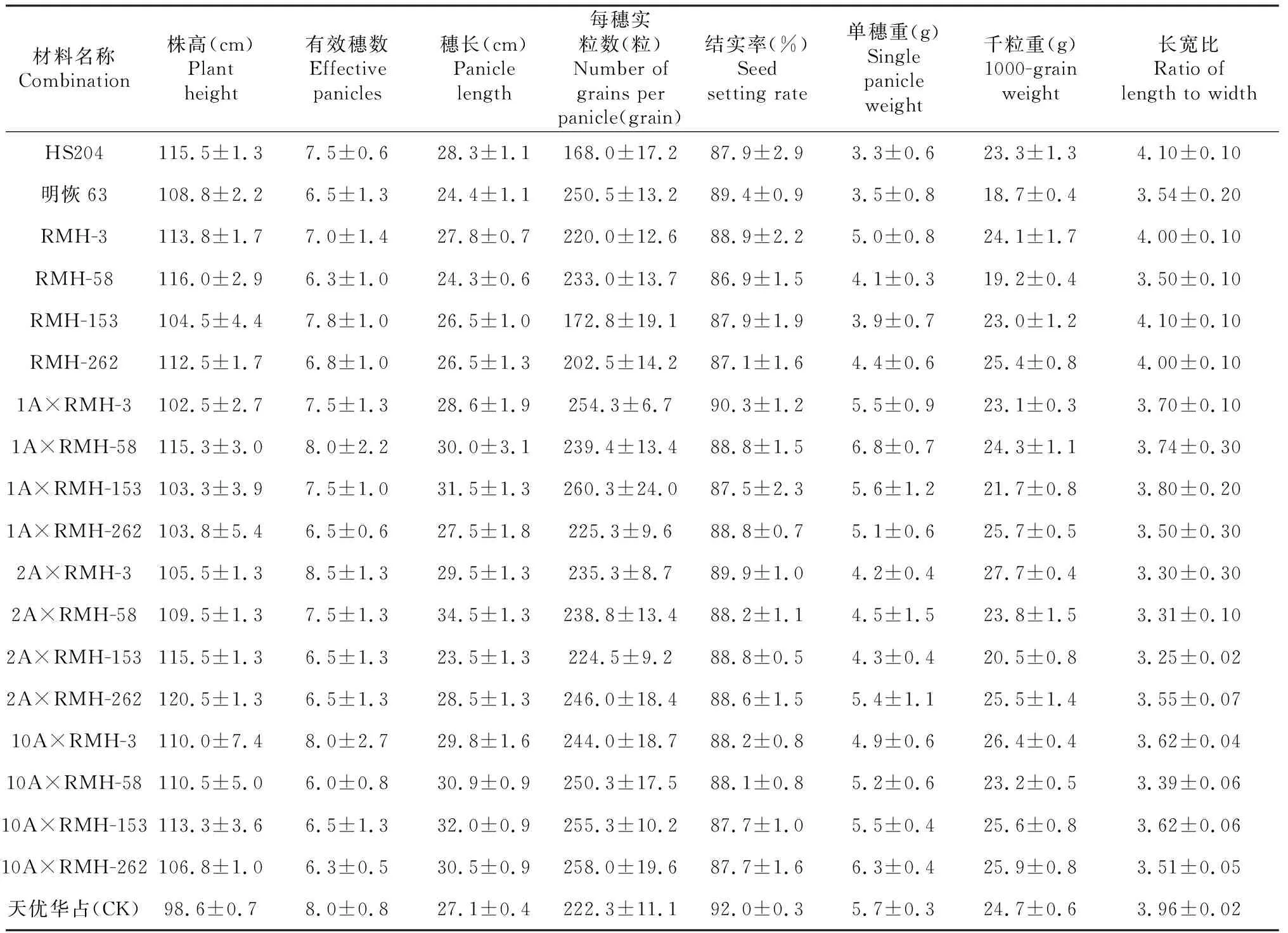
表3 改良株系与杂交配组组合的产量和主要农艺性状表现Table 3 The yield and main agronomic traits of the improved lines and their hybrids

表4 改良株系所配组合对褐飞虱的抗性评价结果Table 4 Resistance of the improved lines and their combinations against BPH
2.4 改良株系配制杂交组合的抗性评价
2018年秋季,对改良株系所配制的12个杂交组合及感虫品种TN1和抗虫品种PTB33进行抗性评价,结果(表4)表明,12个组合中有4个组合表现高抗,平均抗级1.8~1.9;有5个组合表现抗,平均抗级2.7~3.9;有2个组合表现中抗,平均抗级4.0~4.6,有1个组合表现感,平均抗级7.0。说明改良株系组配杂交组合的整体抗性表现良好。
3 讨 论
目前,分子标记辅助选择在低世代仅对目标基因进行选择,无需进行抗性鉴定,具有选育效率高、准确性好、经济和不受环境影响等优点,已广泛应用于水稻抗性育种。如夏明元等[22]选育出含Bph14基因的恢复系R476;李进波等[23]通过回交转育结合分子标记辅助选育,将GD-7的抗稻瘟病基因转育到受体扬稻6号中,获得了10个双基因纯合且农艺性状优良的水稻新品系;兰艳荣等[24]利用分子标记辅助选择改良水稻光温敏核不育系华201S的白叶枯病抗性,获得4个表现良好的抗性株系;阳海宁等[25]采用回交与分子标记辅助选择相结合的方法,获得抗稻白叶枯病基因Xa23和抗稻褐飞虱基因Bph3双抗性基因聚合系144份;闫成业等[25]采用分子标记辅助选择技术与传统育种的杂交、回交相结合,将抗稻瘟病基因Pi9导入R1005中,育成的恢复系含有抗性基因,抗性较父本明显增强。本研究结果与上述研究报道相似,通过分子标记辅助选择获得了褐飞虱抗性较父本明恢63明显增强的恢复系株系。
野生稻是水稻育种的宝贵种质资源。胡杰等[27]研究表明,在水稻中已有30多个抗褐飞虱主效基因被报道,其中有20个基因来源于野生稻,本研究中含有显性基因抗源的供体亲本(母本)HS204也是利用广西普通野生稻培育获得的高抗品系。本研究通过分子标记辅助选育方法将广西本土野生稻的优良抗性基因与恢复系明恢63进行杂交,结合苗期鉴定,在后代中选育出对褐飞虱抗性表现较好的4个改良株系,其农艺性状和产量性状表现优良,配合力强,可作为配制抗褐飞虱杂交组合的遗传资源利用。
在实际生产中,褐飞虱的大暴发多发生在水稻成株期,但部分水稻品种苗期和成株期对褐飞虱的抗性不一致,有些品种是苗期抗虫、成株期感虫,而有些品种是苗期感虫、成株期抗虫[28],本研究还需在目前工作的基础上将获得的抗褐飞虱改良恢复系进行成株期抗性鉴定,并进行持续和广泛的测配,以进一步明确所获得改良恢复系RMH-3、RMH-58、RMH-153和RMH262的褐飞虱抗性表现。
4 结 论
以分子标记辅助选育方法将广西本土野生稻的优良抗褐飞虱基因与恢复系明恢63进行杂交,结合苗期鉴定,在后代中选育出4个改良恢复系RMH-3、RMH-58、RMH-153和RMH262,其褐飞虱抗性表现佳,农艺性状和产量性状表现良好,配合力强,可作为配制抗褐飞虱杂交组合的遗传资源利用。
猜你喜欢
当代水产(2022年7期)2022-09-20
上海农业科技(2022年4期)2022-08-15
热带作物学报(2022年5期)2022-06-06
热带农业科学(2022年5期)2022-06-02
中国饲料(2021年17期)2021-11-02
江西农业学报(2021年8期)2021-09-08
中国蔬菜(2021年3期)2021-03-29
上海师范大学学报·自然科学版(2020年5期)2020-12-18
农村实用技术(2020年9期)2020-12-16
天津农林科技(2020年1期)2020-04-08
