
苏姜猪OLR1 基因多态性及其与肉质性状的关联分析
2020-06-09 08:44任善茂王温慧陶勇
生物技术通报 2020年5期
任善茂 王温慧 陶勇
(江苏农牧科技职业学院,泰州 225300)
血凝素样氧化低密度脂蛋白受体1(Lectin-like oxidized low-density lipoprotein-receptor-l,OLR1) 是一种C 型血凝素家族的氧化型低密度脂蛋白受体,属于Ⅱ型单链跨膜糖蛋白,主要在胎盘、肺、骨髓等组织丰富表达,在心脏、骨骼肌、卵巢等组织中也有少量表达[1-2]。目前,相关研究报道主要集中在OLR1基因多态性与人类心血管疾病和奶牛产奶性状等方面[3-7]。刘旭等[3]采用PCR-LDR 法检测了动脉粥样硬化性脑梗死患者群体OLR1基因3′端G501C 位点多态性,发现病例组CC+GC 基因型、GC 基因型频率显著地高于对照组,从而推测该多态位点可能与动脉粥样硬化性脑梗死相关。Wang 等[5]首次探讨了OLR1基因SNPs 对非白种人迟发性阿尔茨海默病的影响,在中国汉族人群并没有两者之间存在任何显著性差异。Khatib 等[7]通过基因组测序技术在奶牛试验群体中发现了8 个OLR1基因SNPs,其中3′-UTR 区域8 232 bp 处SNP 对脂肪产量和乳脂率有显著影响,AA 基因型个体的OLR1 表 达降低。
苏姜猪是一个通过国家畜禽遗传资源委员会审定的国家级新品种猪,培育过程中采用了现代分子遗传育种技术,其亲本为地方品种姜曲海猪、国外引进品种杜洛克猪[8]。目前,关于苏姜猪的研究主要集中在生产性能、屠宰测定、基因表达等方面[9-11],而有关候选基因多态性与生产性能、肉质性状等指标相关的研究报道较少[12-13]。为了寻找与苏姜猪肉质性状相关的遗传标记,本试验采用PCR-RFLP 技术检测OLR1 基因PstⅠ酶切片断多态性,统计该多态位点的群体遗传参数,分析基因型与肉品指标间的相关关系,以期为分子标记辅助选择技术在苏姜猪持续选育中的应用提供参考依据。
1 材料与方法
1.1 材料
江苏苏姜种猪有限公司提供日龄相近、体重无显著差异的育肥期苏姜猪800 头,公母各半,饲粮营养水平和饲养管理方式一致。无菌采集所有试验用苏姜猪耳组织样后按照KAPA 试剂盒说明书提取组织DNA,采用Scan Drop 100 超微量核酸蛋白测定仪检测DNA 浓度和纯度,稀释后-20℃保存备用。
1.2 方法
1.2.1 肉质性状测定 试验猪体重达到100±5 kg时,每种基因型选取8 头进行屠宰,从左半侧胴体倒数第3-4 胸椎向后采集背最长肌20-30 cm,按照“NY/T 821-2004 猪肌肉品质测定技术规范”测定pH24、失水率、大理石纹、肉色等肉质指标。其中肉色采用5 分制比色板评分法和WSC-S 色差仪法两种方法测定,色差仪测定亮度(L*)、红值(a*)和黄值(b*)。
1.2.2 PCR 扩增 根据郭咪咪等[14]研究成果,由上海生工有限公司合成一对引物,扩增OLR1基因第5 内含子内583 bp 片段。引物序列分别为:上游5′-CGGTG GAAGA GCATT TGT-3′;下 游5′-ACCTT GGGTC AGGCA CTT-3′。PCR 扩增反应体系包括:DNA 模板2.5 μL,2×Taq PCR Mix 12.5 μL,上下游引物(10 μmol/L)各1.0 μL,加dd H2O 至总体积25.0 μL。PCR 反应程序:94℃预变性6 min;94℃变性40 s,53℃退火40 s,72℃延伸50 s,总计35 个循环;72℃延伸10 min,4℃保存。PCR 扩增产物用1.5%琼脂糖凝胶电泳检测,在凝胶成像系统下观察扩增效果。
1.2.3 PCR-RFLP-PstⅠ酶切分析 符合要求的PCR扩增产物用PstⅠ内切酶(赛默飞世尔科技有限公司,中国)进行酶切,酶切反应体系包括1×Buffer 1.5 μL,限制性内切酶0.2 μL(5 U),PCR 产物5 μL,加dd H2O 至15 μL。37℃酶切反应3 h。用1.5%琼脂糖凝胶电泳检测经过限制性内切酶PstⅠ酶切的产物,并在凝胶成像系统下判断基因型。
1.2.4 统计分析
1.2.4.1 分析OLR1基因多态位点的群体遗传参数 参照刘海霞等[15]研究结果计算苏姜猪试验群体纯合度(H0)、杂合度(He)、有效等位基因数(Ne)、多态信息含量(PIC)等OLR1 基因多态位点的群体遗传参数。
纯合度计算公式:

式中H0为基因位点的纯合度,Pi为第i 个等位基因的频率,n 为某个位点的等位基因数。杂合度计算公式:

有效等位基因数计算公式:

多态信息含量计算公式:
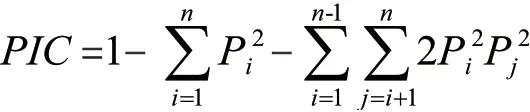
式中n 为等位基因数目,Pi和Pj分别为第i 和第j个等位基因在群体中的频率。
1.2.4.2OLR1基因型与肉质性状间的关联分析 采用SPSS 16.0 统计软件中GLM 过程进行最小二乘方差分析,比较苏姜猪试验群体中OLR1基因型对肉质性状的影响。由于试验群体来自于相同猪场、相同品种,屠宰日龄与体重相近,所以去掉场效应、年龄效应和品种效应,采用简化模型Y=μ+Dl+Rm+elm,式中Y为肉质指标实际测量值,Dl为性别效应,Rm为基因型效应,elm为随机残差效应。
2 结果
2.1 苏姜猪OLR1基因PCR扩增产物
苏姜猪试验群体OLR1基因第5 内含子目标片段的PCR 扩增产物电泳检测结果见图1。图1 显示本次试验中OLR1基因PCR 产物的特异性好,扩增片段的大小与目标片段一致(583 bp),可以直接用来做后续的RFLP 分析。

图1 苏姜猪OLR1 基因PCR 扩增产物
2.2 苏姜猪OLR1基因PCR-RFLP检测分型
苏姜猪试验群体OLR1基因目标片断PCRRFLP-PstⅠ酶切分析电泳检测结果见图2。从图2可见,OLR1基因PCR 产物在苏姜猪试验群体中检测到了PstⅠ酶切多态性,存在CC、CD 和DD 3 种基因型。

图2 苏姜猪OLR1 基因PCR-RFLP-Pst Ⅰ分析结果
2.3 苏姜猪OLR1基因遗传多态性分析
2.3.1 等位基因和基因型频率OLR1基因PCRRFLP-PstⅠ酶切多态位点等位基因和基因型频率的统计结果见表1。由表1 可知,OLR1基因PCRRFLP-PstⅠ多态位点统计在苏姜猪试验群体中的优势基因型为CC,C 为优势等位基因。
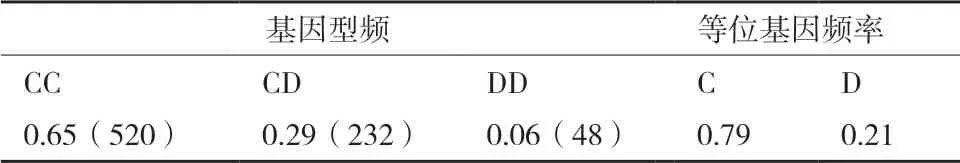
表1 苏姜猪OLR1 基因基因型频率和基因频率
2.3.2 群体遗传纯合度、杂合度等遗传参数分析 苏姜猪试验群体H0、He、Ne及PIC等群体遗传参数统计结果见表2。由表2 可知,苏姜猪试验群体纯合度较高,达到了0.67,多态信息含量为0.27,呈现中度多态。

表2 苏姜猪OLR1 基因多态位点的群体遗传参数
2.4 苏姜猪OLR1基因型与肉质性状的关联分析
苏姜猪OLR1基因PCR-RFLP-PstⅠ酶切多态位点基因型与个体间肉质性状的关联分析结果见表3。由表3 可知,CC 型个体肌肉失水率、大理石纹均显著高于DD 型(P<0.05),CD 和DD 型个体用色差仪测得的黄值b*存在着显著差异(P<0.05),其余指标间的差异均不显著(P>0.05)。
3 讨论
日本学者Swamura 最早在1997 年在牛主动脉内发现了氧化低密度脂蛋白受体-1(OLRl),主要存在于血管等内皮细胞内,能特异性的结合、吞隨、降解氧化低密度脂蛋白[16]。OLRl基因在脾、肾、肺、肝等血管比较丰富的器官中高度表达,而在心脏、卵巢中等器官中的表达量很少[17]。目前对ORL1基因的研究主要集中在人、牛等方面,在猪中研究报道较少[18-21]。Chui 等[18]研究发现人体血液中胆固醇含量随着ORL1 表达量的增加而提高,摄取的游离脂肪酸和脂滴中甘油三酯也随之升高,这些研究结果表明OLR1基因与甘油三酯储存及肥胖产生的关联程度较高。李托等[19]在秦川牛群体中研究发现OLR1基因存在能够被PstI限制性酶识别的g.10497C>A 突变,存在AA、AC 和CC 三种基因型,CC 基因型秦川牛个体的眼肌面积和眼肌深度均显著地高于其他两种基因型个体。刘春伟等[20]克隆测序后分析得到猪OLR1基因编码区全长为825 bp(GenBank 序列号EU122440),编码273 个氨基酸,猪OLR1基因与牛同源性高达84%,其次与人、大鼠、小鼠的同源性分别是79%、51%和27%。郭咪咪等[14]通过RT-PCR 分析发现猪OLR1基因在不同组织间的表达差异很大(心脏内表达量最高),在背脂中表达呈现出明显的品种差异。

表3 苏姜猪OLR1 基因不同基因型与肉质性状间的关联分析
本研究在苏姜猪试验群体中发现OLR1基因PstⅠ酶切位点多态性,测得C、D 两个等位基因,CC、CD、DD 三种基因型。根据郭咪咪等[21]研究结果,该多态位点为位于OLR1基因第5 内含子内的 C52T 突变,因此该多态位点并不会导致所编辑的氨基酸突变,但是否会影响OLR1基因的表达还有待深入研究。该多态位点在苏姜猪试验群体的多态信息含量(PIC)为0.27,呈现出中度多态,由此可见苏姜猪群体具备进一步选育的潜力。郭咪咪等[21]在长白猪、大白猪、梅山猪等中外试验猪群体中均未检测到酶切基因型的纯合个体,不同基因型在大白×梅山F2 代杂交商品猪的肉色指标间也不存在显著或极显著的差异。但在本研究中却发现OLR1 基因不同基因型个体在失水率、大理石纹、色差仪测得的肉色黄值b*指标间均存在显著差异。这些研究结论的差异可能与苏姜猪的遗传背景有关。苏姜猪是以姜曲海猪、枫泾猪、杜洛克猪为亲本培育而成的新品种猪,分别含有姜曲海猪血统18.75%、枫泾猪血统18.75%和杜洛克猪血统62.5%[22],因此杜洛克猪是否是导致苏姜猪OLR1 基因酶切基因型纯合个体出现的原因还有待验证。中国地方猪种肉质优良,大理石纹丰富,肌内脂肪含量高达3%以上,有的甚至达到6%-7%[23],苏姜猪也具有我国地方猪种姜曲海猪肉质优良的特性[24]。因此,从本研究中OLR1 基因与肉质性状关联性分析得出,目前可以通过提高苏姜猪群体中CC 基因型个体的频率来增加肌肉大理石纹含量,从而达到改善肉质的目的,这也为苏姜猪肉质性状的分子辅助选育提供了参考 依据。
4 结论
本试验在苏姜猪试验群体OLR1基因第5 内含子内检测到一个PCR-RFLP-PstⅠ多态位点,存在C、D 两个等位基因,CC、CD、DD 三种基因型,其中C 为优势等位基因,CC 为优势基因型。关联分析表明,该多态位点不同基因型与失水率、大理石纹、肉色等肉质指标间差异显著。因此,本试验检测到的PCR-RFLP-PstⅠ多态位点可作为肉质性状候选基因在苏姜猪的持续选育中加以应用。
猜你喜欢
今日农业(2022年2期)2022-11-16
美食(2022年4期)2022-04-16
智慧健康(2021年17期)2021-07-30
美食(2020年5期)2020-06-01
新世纪智能(高一语文)(2020年12期)2020-06-01
中国产前诊断杂志(电子版)(2020年1期)2020-05-21
遵义医科大学学报(2020年6期)2020-02-05
科技视界(2016年27期)2017-03-14
现代检验医学杂志(2016年5期)2016-08-20
中国医药导报(2015年27期)2015-02-28
