
共表达HAC1 和分子伴侣基因对甘露聚糖酶在毕赤 酵母中表达的影响
2020-06-09 08:45闵琪高子涵姚银张华山熊海容张莉
生物技术通报 2020年5期
闵琪 高子涵 姚银 张华山 熊海容 张莉
(1. 中南民族大学生命科学学院,武汉 430074;2 湖北工业大学发酵工程省部共建教育部重点实验室,武汉 430068)
β-甘露聚糖酶(β-1,4-mannanase,EC 3.2.1.78)是降解甘露聚糖的关键酶,属于内切水解酶,能降解含 β-1,4-甘露糖苷键的甘露寡糖和甘露多 糖[1-2]。β-甘露聚糖酶在饲料、食品、医药、石油开采等方面具有广泛应用前景[3-4]。然而目前甘露聚糖酶的工业生产成本较高,使其在实际生产应用中受到一定制约。利用基因工程的方法提高甘露聚糖酶的产量在其工业化生产中具有十分重要的意义,而共表达分子伴侣是提高外源蛋白表达的有效策略 之一[5-6]。
当蛋白高水平表达时,未折叠的蛋白质容易在内质网中错误折叠和累积,内质网主要通过UPR 调控这一现象,UPR 激活调控因子HAC1 的表达,可促进 UPR 相关蛋白质及分子伴侣的高水平表达,介导蛋白质的正确折叠分泌,及错误折叠蛋白在内质网中的相关降解(ERAD)[7-8]。大量研究表明分子伴侣对外源蛋白的分泌表达有着重要意义,其主要作用是辅助蛋白质的正确折叠,减少由于蛋白质错误折叠而导致的降解,从而提高蛋白质的分泌表 达[7,9]。ERO1 在内质网中氧化还原态的蛋白二硫键异构酶PDI 成氧化态,PDI 的氧化活性帮助蛋白形成正确的折叠结构而分泌到胞外,对内质网腔内蛋白质二硫键的形成具有促进作用,且ERO1、PDI两种分子伴侣都负责内质网氧化蛋白的折叠[10-11]。分子伴侣如PDI1,也属于蛋白二硫化物异构酶,是内质网腔中多功能分子伴侣[12];CPR5 亲环蛋白,参与蛋白质翻译后修饰,维持细胞内金属离子的内稳定[13]。BiP / Kar2p,重链结合蛋白,防止蛋白质分子内和分子间的相互作用而导致的错误折叠或聚集,并与错误折叠或未折叠的蛋白质结合,并阻止它们离开内质网[14]。Huang 等[15]通过将HAC1 在分泌α-淀粉酶的毕赤酵母重组菌胞内共表达,使重组菌的α-淀粉酶在摇瓶水平上的酶活提高约2 倍。Zhang 等[16]将内质网中的KAR2(BiP)与PDI 两种分子伴侣组合,并同时在分泌G-CSF 蛋白的毕赤酵母重组菌胞内共表达,结果重组菌在摇瓶水平分泌的G-CSF 蛋白总量增加6.5 倍。Zhang 等[17]通过在分泌表达β-葡萄糖苷酶的毕赤酵母重组菌中胞内共表达PDI,共表达PDI 的重组菌在3 L 发酵罐中诱导发酵时,其发酵上清液的酶活提高46%,同时发酵上清液中β-葡萄糖苷酶的比活力提高54%。
本研究将HAC1 及5 种内质网折叠相关分子伴侣分别在分泌表达ManA 的重组菌中胞内共表达,并进一步将HAC1、ERO1、PDI进行两基因或三基因组合后再进行共表达,以研究HAC1 及不同分子伴侣对ManA 在毕赤酵母GS115 中分泌表达的影响。
1 材料与方法
1.1 材料
1.1.1 菌种与质粒 分泌表达ManA 的重组菌GS115/pPIC9K-ManA 由实验室构建并保藏(ManA基因编号为Accession No. KJ806637[18])。表达载体pPICZA、巴斯德毕赤酵母GS115、大肠杆菌Top10均由实验室保藏。
1.1.2 主要试剂与仪器 限制性内切酶、Prime STAR Max DNA 聚合酶、T4 DNA 连接酶等工具酶、DNA Marker、TaKaRa Bradford Protein Assay Kit 均购自宝生物工程(大连)有限公司;Prestained protein ladder 购自赛默飞公司。质粒小量提取试剂盒、DNA 凝胶回收试剂盒、PCR 清洁回收试剂盒等购自上海AxyPrep 公司;SDS-PAGE 凝胶快速配置试剂盒购于碧云天公司;底物角豆胶购于Sigma 公司;Tryptone和Yeast Extract 购自于Oxoid 公司;生物素、无氨基酵母氮源YNB、山梨醇和琼脂购自Biosharp 公司;博来霉素Zeocin 购自Invitrogen 公司;其他试剂均为国产或进口分析纯。
1.1.3 培养基
LBL(1 L):10 g 蛋白胨,5 g NaCl,5 g 酵母提取物(使用时按需要加入zeocin,根据Invitrogen 使用说明书)
YPD(1 L):10 g 酵母提取物,20 g 蛋白胨,20 g 葡萄糖(使用时按需要加入zeocin,根据Invitrogen使用说明书)
BMGY(1 L):10 g 酵母提取物,20 g 蛋白胨,13.4 gYNB,1 mL 500×生物素(4×10-4g/L),100 mL 1 mol/L pH 6.0 磷酸钾缓冲溶液,1%甘油(根据毕赤酵母表达手册配置)
BMMY(1 L):10 g 酵母提取物,20 g 蛋白胨,13.4 gYNB,1 mL 500×生物素(4×10-4g/L),100 mL 1 mol/L pH 6.0 磷酸钾缓冲溶液,甲醇根据培养所需添加(根据毕赤酵母表达手册配置)。
1.2 方法
1.2.1 HAC1 与5 种分子伴侣胞内表达重组质粒及对应重组菌的构建
1.2.1.1 六种重组质粒的构建 HAC1 与5 种分子伴侣基因序列NCBI 数据库编号分别是CPR5(XM_002489874.1,669 bp);HAC1(XM_002489994.1,996 bp);PDI1(XP_002489806.1,1 110 bp);PDI(AJ302014.1,1 554 bp);ERO1(XM_002489600.1,1 584 bp);BiP(XM_002490982.1,2 037 bp)。 以GS115 基因组为模板,分别扩增6 种基因,各PCR扩增引物如表1,下划线为酶切位点。将PCR 所得的产物与载体pPICZA 分别双酶切、连接并转化大肠杆菌Top10,挑取阳性转化子培养后提取质粒送至武汉擎科生物技术有限公司进行测序验证。
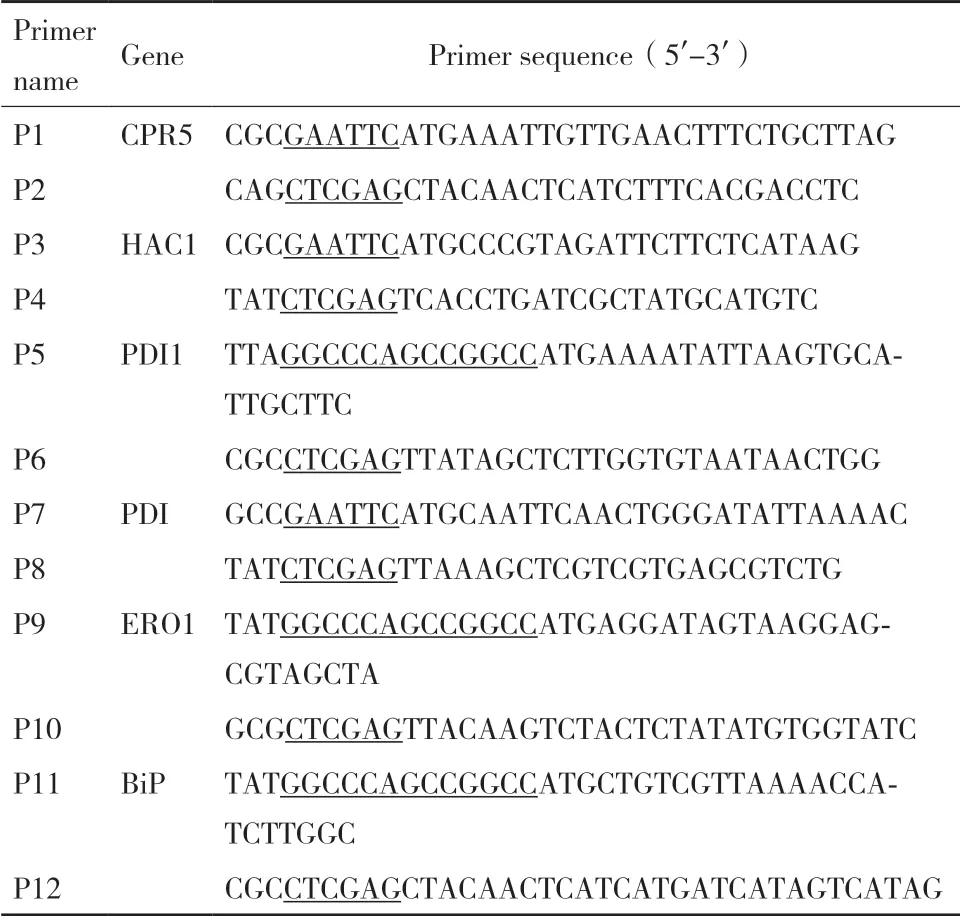
表1 PCR 扩增引物序列
1.2.1.2 六种重组菌的构建 采用SacI 分别线性化上述构建好的重组质粒,电转化至已制备成感受态的宿主菌GS115/pPIC9K-ManA 中,涂布于YPDZ博来霉素抗性筛选平板上,筛选获得阳性转化子,并分别命名为GS115/ManA-CPR5、GS115/ManAHAC1、GS115/ManA-PDI1、GS115/ManA-PDI、GS115/ManA-ERO1、GS115/ManA-BiP。
1.2.2 六种重组菌的诱导表达及相关分析
1.2.1 试验地点 试验设在凉山州普格县五道箐乡采洛洛博村进行,海拔2 000 m,土质为黄棕壤,试验地肥力中等,前茬作物为玉米,播种前拖拉机旋耕整地耙平。
1.2.2.1 六种重组菌的摇瓶发酵 将各重组菌接种于10 mL BMGY 培养基中,30℃、250 r/min 培养24 h,离心收集适量菌体,重悬于25 mL BMMY 培养基中,控制起始OD600值约为1.0,在30℃、250 r/min 条件下培养至192 h,每隔24 h 取样0.25 mL 并补加1%甲醇,检测各样品的OD600值(通过测量600 nm 处的OD 值来确定酵母细胞生长状况)和酶活力,以GS115/pPIC9K-ManA 作为对照。所有实验数据均重复3 次,并通过GraphPad Prism 5 软件对数据进行显著性分析。
甘露聚糖酶酶活力采用DNS 法[19]测定,将收取的样品离心并收集上清,以pH6.0 的Na2HPO4-柠檬酸缓冲液配置的0.5%角豆胶为底物,酶活力单位定义为在其最适温度75℃和最适pH 6.0 条件下,1 min 内水解甘露聚糖底物产生1 μmol 甘露糖所需要的甘露聚糖酶的量。
1.2.2.2 六种重组菌胞内滞留酶活检测 摇瓶发酵至168 h 时取等量菌体,10 000 r/min 离心,弃上清,用等体积预冷的pH 6.0 的Na2HPO4-柠檬酸缓冲液重悬,重复洗涤2 次。再用等量的缓冲液重悬,向每管中加入0.73 g 酸性玻璃珠,振荡器上处理30 min,离心去除细胞碎片,收集上清,测定胞内甘露聚糖酶酶活力。所有实验数据均重复3 次。
1.2.2.3 六种重组菌发酵上清液蛋白浓度的测定及SDS-PAGE 电泳 摇瓶发酵至168 h 时,取等体积的发酵上清液进行SDS-PAGE 电泳分析,并根据TaKaRa Bradford Protein Assay Kit 使用说明检测发酵上清液总蛋白含量。所有实验数据均重复3 次。
1.2.3 HAC1、ERO1、PDI 组合重组质粒及对应重组菌的构建
1.2.3.1 四种组合重组质粒的构建 胞内共表达HAC1、ERO1 和PDI 的重组菌发酵上清液中的ManA酶活力分别有不同程度的提升,因此选择HAC1、ERO1 和PDI 构 建4 种 组 合 质 粒pPICZA/HAC1-ERO1、pPICZA/HAC1-PDI、pPICZA/ERO1-PDI、pPICZA/HAC1-ERO1-PDI。首先采用重叠延伸PCR将HAC1、ERO1 和PDI 基因中的BglII 和BamH I位点在不影响翻译后蛋白序列的条件下定点突变,去除各基因中的BglII 和BamH I 位点,重叠延伸PCR 引物见表2。然后根据同尾酶法构建各两基因组合重组质粒,以pPICZA/HAC1-ERO1 为例,流程图如图1 所示。在已构建好的pPICZA/HAC1-ERO1的基础上进一步构建pPICZA/HAC1-ERO1-PDI,方法同上。

表2 重叠延伸引物序列
1.2.3.2 四种重组菌的构建 将4 种组合重组质粒分别电转化至已制备成感受态的宿主菌GS115/pPIC9K-ManA 中,筛选阳性转化子,并分别命名为GS115/ManA-HAC1-ERO1、GS115/ManA-HAC1-PDI、GS115/ManA-ERO1-PDI、GS115/ManA-HAC1-ERO1-PDI。
1.2.4 四种重组菌的诱导表达及相关分析 各重组菌摇瓶发酵及相关分析实验同1.2.2。
2 结果
2.1 HAC1与5种分子伴侣重组质粒及对应重组菌的构建
以提取的GS115 基因组为模板,PCR 扩增获得6 种 基 因,CPR5(669 bp),HAC1(996 bp),PDI1(1 110 bp),PDI(1 554 bp),ERO1(1 584 bp),BiP(2 037 bp),扩增结果如图2,大小与预期一致。将目的基因分别与载体pPICZA 连接获得6 种重组质粒,测序结果表明6 种重组质粒均构建成功。然后将各重组质粒电转化至GS115/pPIC9K-ManA 中,筛选获得阳性转化子。
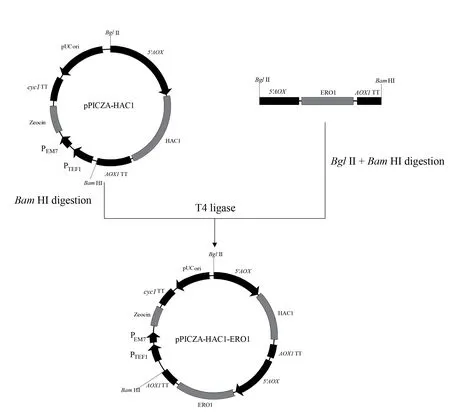
图1 pPICZA/HAC1-ERO1 质粒构建示意图

图2 六种基因的PCR 产物电泳图
2.2 六种重组菌的诱导表达及相关分析
2.2.1 六种重组菌的生长曲线及酶活曲线 各重组菌发酵生长曲线及酶活曲线如图3 所示,从图3-A可以看出各重组菌的生长趋势一致,表明HAC1 及5 种分子伴侣的胞内共表达不会影响重组菌的正常生长。从图3-B 中发现各重组菌摇瓶发酵至168 h 时,上清的酶活力达到最高,其中GS115/ManA-HAC1、GS115/ManA-ERO1、GS115/ManA-PDI 的酶活水平相比GS115/ManA 分别提高26%、15%、20%,分别达到1 014 U/mL、925 U/mL、965 U/mL。
2.2.2 六种重组菌酶活力的比较 摇瓶发酵至168 h时,检测各重组菌上清的酶活及胞内滞留酶活,并计算各重组菌的总酶活见图4。重组菌GS115/ManAHAC1、GS115/ManA-ERO1、GS115/ManA-PDI 分 泌至上清液的酶活分别提高26%、15%、20%,而GS115/ManA-HAC1 胞内滞留的酶活力下降34%,且GS115/ManA-ERO1 和GS115/ManA-PDI 胞内滞 留 酶活力也略有下降,同时3 种重组菌发酵产生的总酶活分别提高了22%、13%、18%;重组菌GS115/ManA-PDI1 和GS115/ManA-CPR5 发酵上清液的酶活及发酵产生的总酶活力相对GS115/ManA 无明显变化;而重组菌GS115/ManA-BiP 发酵上清液的酶活下降了22%,胞内滞留酶活反而上升了42%,总酶活力下降17%。
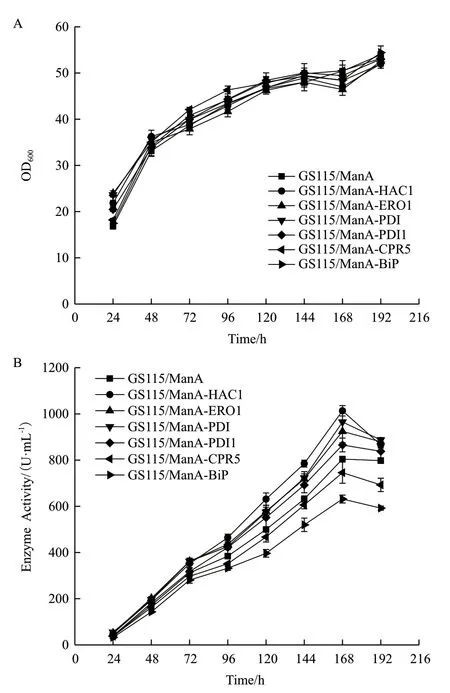
图3 六种重组菌生长曲线(A)及酶活力曲线(B)
2.2.3 六种重组菌发酵上清液蛋白浓度的测定及SDS-PAGE 电泳 摇瓶发酵至168 h 时,对各重组菌上清液进行SDS-PAGE 电泳(图5),测定各重组菌上清液总蛋白含量并计算出比活力(表3),结果发现胞内共表达HAC1、ERO1、PDI 基因的重组菌,发酵上清液中的比活力分别提高了65%、55%、44%,说明上清液中外源蛋白ManA 的纯度相对 提高。
2.3 HAC1、ERO1、PDI组合重组质粒及对应重组菌的构建
将HAC1 ERO1 和PDI 进行两基因或三基因组合,按照图1 流程构建4 种组合重组质粒,使用BglII 与BamH I 进行双酶切鉴定结果如图6,双酶切鉴定大小与预期一致。将各组合重组质粒电转化至GS115/pPIC9K-ManA 中,筛选各重组菌阳性转化子进行后续实验。
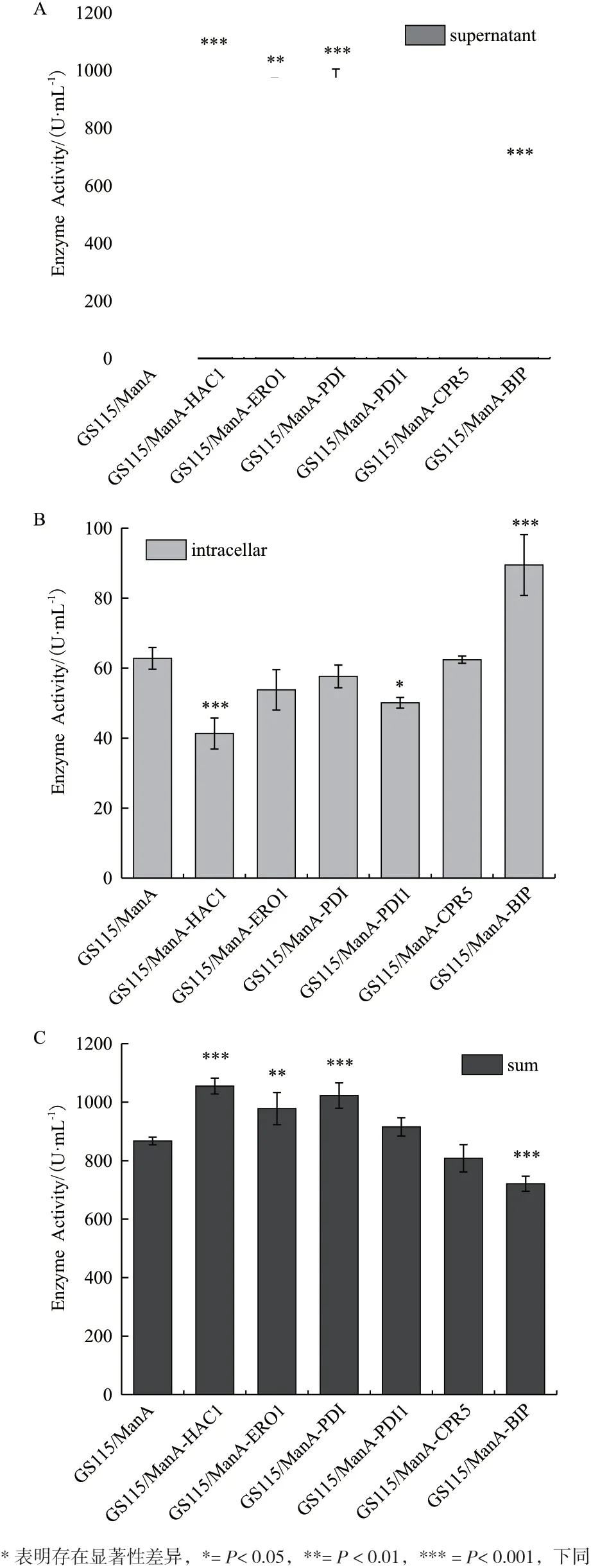
图4 六种重组菌酶活力分析(168 h)
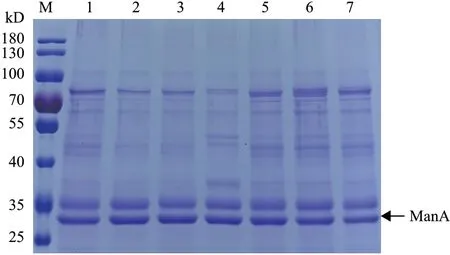
图5 六种重组菌发酵上清液的SDS-PAGE 分析(168 h)
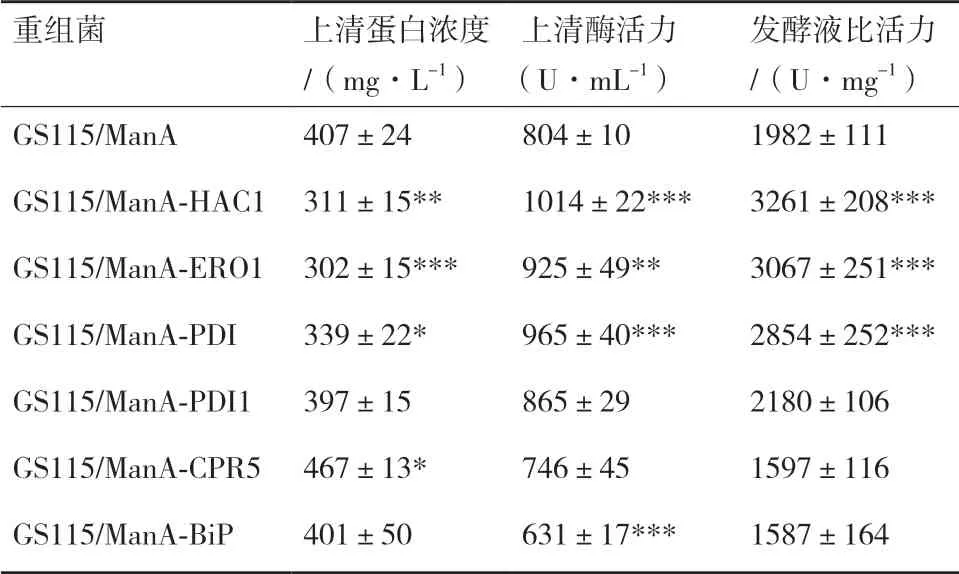
表3 六种重组菌ManA 发酵上清液蛋白含量及比活力 分析(168 h)
图6 四种重组质粒双酶切鉴定
2.4 四种重组菌诱导表达及相关分析
2.4.1 四种重组菌的生长曲线及酶活曲线 各重组菌生长曲线与酶活曲线见图7。图7-A 显示各重组菌生长趋势无明显差异,表明各重组菌均生长正常。从图7-B 可以看出,各重组菌摇瓶发酵至168 h 时,GS115/ManA-HAC1-PDI 上清 酶 活 比GS115/ManA 提高10%,达到884 U/mL,而其它重组菌上清酶活力与GS115/ManA 相比均无明显变化。
图7 四种重组菌生长曲线(A)及酶活曲线(B)
2.4.2 四种重组菌酶活力的比较 摇瓶发酵至168 h 时,测定各重组菌上清酶活及胞内滞留酶活,并计算各重组菌的总酶活力见图8。从图8 可以看出,GS115/ManA-HAC1-PDI 发酵上清液酶活提高10%,且发酵产生的总酶活力提高10%,而其它3 种重组菌的上清酶活、胞内滞留酶活及总酶活都无明显变化。基于各重组菌的酶活力比较,GS115/ManAHAC1-PDI 对ManA 的分泌表达有一定促进作用,但相 较 于GS115/ManA-HAC1、GS115/ManA-ERO1 和GS115/ManA-PDI 却并没有进一步的提升。
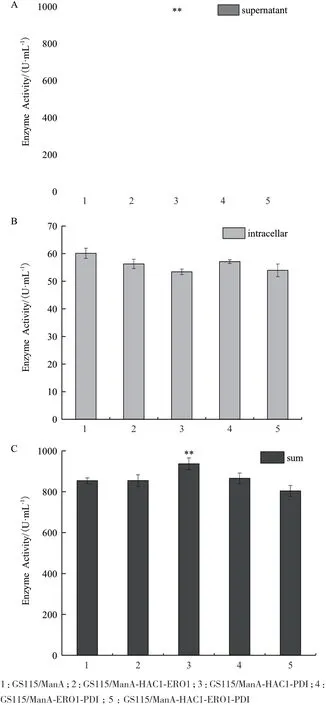
图8 四种重组菌酶活力分析(168 h)
2.4.3 四种重组菌发酵上清液蛋白浓度的测定及比活力计算 摇瓶发酵至168 h 时,检测各重组菌发酵上清液的总蛋白含量并计算比活力,结果见表4,同时对各上清液进行SDS-PAGE 电泳分析见图9。对 比GS115/ManA,GS115/ManA-HAC1-PDI 比活 力提高39%,GS115/ManA-HAC1-ERO1-PDI 比活力提高27%,而其它3 种重组菌发酵上清液的比活力无明显变化。但GS115/ManA-HAC1-PDI 发酵上清液的ManA 比活力相比 GS115/ManA-HAC1、GS115/ManAERO1、GS115/ManA-PDI 却没有进一步提高。

表4 四种重组菌ManA 发酵上清液蛋白含量及比活力分析(168 h)
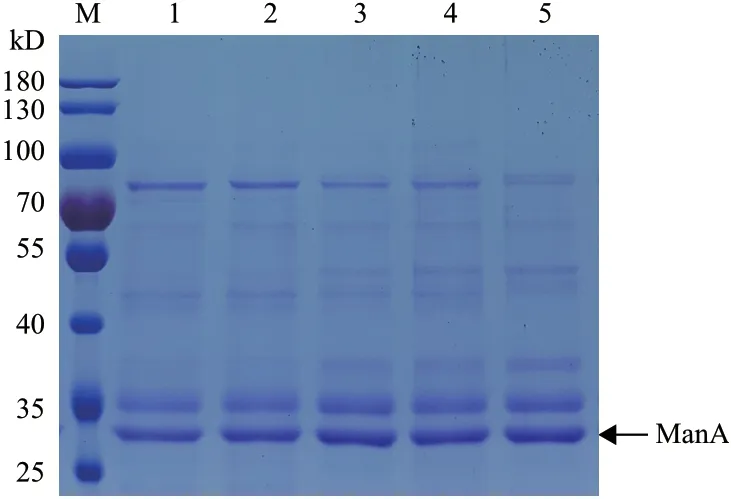
图9 四种重组菌发酵上清液的SDS-PAGE 分析(168 h)
3 讨论
外源蛋白的大量表达可能会使ER 折叠和分泌外源蛋白质的能力造成负担,进而错误折叠或未折叠的蛋白在ER 中大量积聚,导致未折叠蛋白应答(UPR)途径的激活,该途径通过诱导参与蛋白折叠的分子伴侣和ER 相关降解(ERAD)途径的基因来减轻ER 的压力[7-8]。分泌表达ManA 的重组菌胞内共表达HAC1、ERO1 和PDI 时,各重组菌发酵上清液的酶活水平分别提高26%、15%和20%,且胞内滞留酶活分别下降,表明了HAC1 及分子伴侣ERO1和PDI 的共表达对甘露聚糖酶ManA 的正确折叠及分泌表达具有一定的促进效果,说明其中HAC1 的表达,可能是其介导的UPR 途径,辅助外源蛋白ManA 的正确折叠及分泌,减少由于ManA 错误折叠而导致的ER 相关降解途径(ERAD)的降解,从而提高ManA 的分泌表达[7-8]。而分子伴侣如ERO1,PDI 的共表达,主要是直接或间接通过其蛋白的分子伴侣活性介导未折叠的蛋白质正确折叠,及错误折叠蛋白的再折叠来辅助表达ManA[5]。在分泌表达ManA 的重组菌胞内共表达PDI1、CPR5 和BiP 时,各重组菌发酵液的酶活力没有明显提高,表明并非所有的内质网分子伴侣都能辅助ManA 的正确折叠及分泌表达。对于HAC1 与分子伴侣组合并在分泌表达ManA 的重组菌,其分泌表达ManA 的酶活力没有进一步提升,研究发现外源蛋白的正确折叠与不同的分子伴侣,或者不同的分子伴侣组合之间没有特别对应的规律。已报道的研究中,不同的分子伴侣的组合协同表达外源蛋白,有时比单独一种分子伴侣与外源蛋白共表达的效果要更明显[16,20],但也有不同分子伴侣的组合与外源蛋白共表达时,效果反而不明显的现象[21]。分子伴侣相互之间的影响仍有待进一步研究,针对分子伴侣对酵母表达系统进行优化,应该根据目的蛋白的分子结构特征而区别对待,构建相对最适合的表达工程菌[22]。
在胞内共表达HAC1、ERO1 和PDI 后,各重组菌ManA 发酵上清液的比活力分别提高了65%、55%和44%,表明ManA 发酵上清液的质量也相应得到提高,对工业上生产甘露聚糖酶的过程中简化生产流程以及降低生产成本有着一定的实际意义,但HAC1 及分子伴侣对毕赤酵母分泌表达上清中ManA 的比活力的变化还需要进一步纯化论证。在提高甘露聚糖酶表达量的相关研究中,其它研究策略如改变启动子[23]、信号肽[24]等,虽然对酶活的提升都有一定作用,但同时也可能会导致胞内滞留酶活的积累,增加内质网蛋白折叠的压力,若同时共表达HAC1 及分子伴侣ERO1、PDI 等基因,能协同分泌表达甘露聚糖酶,从而构建高效分泌表达甘露聚糖酶的工程菌。
蛋白质分泌的限制可能是由低折叠率造成,原因可能是内质网相关蛋白折叠分子伴侣的表达水平不足[25]。已报道的研究中发现,HAC1 或者分子伴侣基因以多拷贝的形式在毕赤酵母重组菌胞内共表达可以进一步提高其重组菌外源蛋白的分泌产量,如Huang 等[15]在分泌表达不同拷贝α-淀粉酶基因的毕赤酵母重组菌胞内共表达HAC1,并随着HAC1拷贝数在一定范围的增加,分泌表达不同拷贝α-淀粉酶基因的重组菌在摇瓶发酵水平分泌表达α-淀粉酶酶活都不断提高,其中分泌表达12 拷贝α-淀粉酶基因,并在胞内共表达6 拷贝HAC1的重组菌酶活达到最大值,相比分泌表达12 拷贝α-淀粉酶基因,并在胞内共表达1拷贝HAC1的重组菌提高了约1倍。故本研究中的HAC1 及分子伴侣ERO1、PDI 还可以进一步研究其多拷贝基因在分泌表达不同拷贝ManA的毕赤酵母重组菌胞内共表达,从而构建高效分泌表达ManA 的工程菌。
4 结论
本研究在工程菌GS115/pPIC9K-ManA 的基础上,分别同时共表达HAC1、ERO1、PDI,各重组菌在摇瓶发酵水平,发酵上清液的ManA 酶活力都有所提高,其中整合HAC1 基因的重组菌最高提高了26%。进一步将HAC1、ERO1、PDI进行两基因或三基因组合后再进行共表达,但其重组菌发酵上清液酶活力并没有进一步提升。
猜你喜欢
安徽农业科学(2022年1期)2022-02-14
昆明医科大学学报(2020年12期)2021-01-26
海峡姐妹(2020年6期)2020-07-25
河南化工(2020年6期)2020-07-20
海峡姐妹(2019年12期)2020-01-14
食品安全导刊(2019年27期)2019-12-09
名城绘(2019年4期)2019-10-21
海峡姐妹(2019年9期)2019-10-08
安徽化工(2018年6期)2019-01-11
好日子(2018年9期)2018-10-12
