
多鳞鱚vhl基因克隆及其低氧胁迫下的表达变化
2020-07-07 09:33林星桦潘炎杨陈芳圆黄志棚黄洋杜涛朱春华李广丽田昌绪
南方农业学报 2020年5期
林星桦 潘炎杨 陈芳圆 黄志棚 黄洋 杜涛 朱春华 李广丽 田昌绪


摘要:【目的】克隆多鱗鱚林岛综合征基因(vhl)并分析其在低氧胁迫下的表达变化,为揭示多鳞鱚应对低氧的适应机制提供研究资料,也为今后开展多鳞鱚新品种选育提供候选基因。【方法】采用PCR克隆多鳞鱚vhl基因cDNA全长序列,利用EditSeq、SMART、Signal IP 4.1及ClustalX等在线软件进行生物信息学分析;并以半定量PCR检测多鳞鱚vhl基因在不同组织中的表达情况,采用实时荧光定量PCR分析低氧胁迫对多鳞鱚vhl基因在鳃组织和心脏中表达的影响。【结果】多鳞鱚vhl基因cDNA序列全长727 bp,包括120 bp的5'端非编码区(5'-TUR)、106 bp的3'端非编码区(3'-TUR)和504 bp的开放阅读框(ORF),共编码167个氨基酸残基,编码蛋白存在4个结构域,即Pfam:VHL(第12~93位氨基酸)、Pfam:VHL_C(第5~22、31~39和98~153位氨基酸)、ParB结构域(第100~162位氨基酸)和SOCS_box结构域(第104~140位氨基酸)。多鳞鱚vhl氨基酸序列与盲曹鱼vhl氨基酸序列的同源性最高(81%),但系统发育进化分析结果显示多鳞鱚与攀鲈的亲缘关系最近。多鳞鱚vhl基因在不同组织中均有表达,其中以鳃组织、卵巢和精巢的表达量较高,其次是心脏和肝脏,在肌肉和脑组织的表达量较低。经低氧胁迫后,多鳞鱚vhl基因在鳃组织中的相对表达量显著上调(P<0.05,下同);在心脏中表现为随低氧胁迫时间的延长显著上调,恢复氧气4 h后vhl基因的相对表达量虽然呈显著下调趋势,但仍显著高于正常水平。【结论】多鳞鱚vhl基因编码蛋白存在Pfam:VHL、Pfam:VHL_C、ParB和SOCS_box等4个结构域,且低氧胁迫前后其表达量存在显著差异,即多鳞鱚vhl基因在低氧应答信号通路中扮演着重要角色。
关键词: 多鳞鱚;vhl基因;低氧胁迫;表达变化
中图分类号: S965.399 文献标志码: A 文章编号:2095-1191(2020)05-1201-08
Abstract:【Objective】The Von Hippel-Lindau syndrome gene(vhl) of the Sillago sihama was cloned and its expression changes after hypoxia stress treatment were analyzed, which represented the insights into the underlying adaptation mechanisms of hypoxia stress and provided candidate genes for breed selection of S. sihama. 【Method】cDNA sequence of vhl gene from S. sihama was cloned by PCR. Bioinformatics analysis on the sequence was conducted by online softwares such as EditSeq,SMART, Signal IP 4.1 and ClustalX, expression of vhl gene in different tissues were detected by semiquantitative PCR, and effects of hypoxia stress on expression of vhl gene in gill and heart tissues were studied by real-time fluorescence quantitative PCR. 【Result】The vhl cDNA sequence was 727 bp in length, including a 5'-end noncoding region of 120 bp, a 3'-end noncoding region of 106 bp and an open reading frame(ORF) of 504 bp encoding a polypeptide of 167 amino acids.Four domains of the vhl protein were found, namely Pfam:VHL amino acids 12 to 93, Pfam:VHL_C (amino acids 5 to 22, 31 to 39 and 98 to 153), and ParB domain(amino acid 100 to 162) and SOCS_box domain (amino acids 104 to 140). Multi-sequence alignment analysis of the S. sihama vhl gene showed the highest(81%) homology with Lates calcarifer. Cluster analysis showed that the relationship between S. sihama and Anabas testudineus was the closest. vhl gene expressed in different tissues of S. sihama. Its expression in the gill was significantly up-regulated after hypoxia stress(P<0.05, the same below), the expression of vhl gene in heart significantly up-regulated as hypoxia stress prolonged, and significantly down-regulated 4 h after reoxygen treatment, bur is was still significantly higher than normal level. 【Conclusion】There are four domains in vhl gene encoded protein of S. sihama, namely Pfam:VHL, Pfam: VHL_C, ParB and SOCS_box. The expression of vhl gene before and after hypoxia stress is significantly different, indicating that vhl gene plays an important role in the hypoxia response pathway in S. sihama.
Key words: Sillago sihama; vhl gene; hypoxia stress; expression variation
Foundation item: Scientific Research Project of Guangdong(2015A020209163); College Featured Innovation Pro-ject of Guangdong(2019KTSCX060); College Students Innovation and Entrepreneurship Project of Guangdong(CXXL 2018048, CXXL2019138)
0 引言
【研究意义】缺氧会影响鱼类的生长、繁殖及行为活动,是影响大多数动物有氧代谢与存活的重要环境因素,已成为阻碍水产养殖业健康发展的限制性因子(Pollock et al.,2007;Roberts et al.,2011;武晓会等,2018)。低氧诱导因子(Hypoxia inducible factor,HIF)介导的低氧信号通路是细胞产生低氧应答的核心信号通路之一(李福祥等,2004;Dunwoo-die,2009;肖武汉,2014)。在低氧信号通路中,林岛综合征基因(Von hippel-lindau tumor suppressor,vhl)发挥着重要作用,其介导HIF-1α的表达调控,以确保生物体能及时启动低氧应答。在常氧条件下,vhl基因介导HIF-1α降解,而抑制其下游基因表达;但在低氧条件下,vhl无法利用氧分子使HIF-1α降解,进而引发一系列低氧应答反应(Dunwoodie,2009;肖武汉,2014)。因此,加强vhl基因研究对揭示动物机体的低氧适应机制具有重要意义。【前人研究进展】已有研究表明,在体细胞中肿瘤抑制因子pVHL能与Elongin B和Elongin C结合形成VBC泛素连接酶复合体而具有E3泛素酶蛋白活性,促使羟基化的HIF-1α泛素化并降解(肖武汉,2014)。在常氧条件下,vhl与脯氨酸羟化酶(Prolyl hydroxylase,PHD)结合利用氧气中的氧分子使HIF-1α羟基化、泛素化并降解,限制HIF-1α与HIF-1β结合,从而抑制其下游基因的表达;而在低氧条件下,由于氧气含量下降,vhl无法利用氧分子使HIF-1α降解,HIF-1α与HIF-1β正常结合,激活血管内皮生长因子(Vascular endothelial growth factor,VEGF)和促红细胞生成素(Erythropoietin,EPO)等下游基因表达(Kaelin,2007),引起红细胞增加、血管扩张等低氧应答反应。在小鼠肝脏星状细胞中,vhl蛋白水平下降导致HIF-1α积累并激活VEGF表达,参与肝脏血管生成(Ankoma-Sey et al.,2000;Corpechot et al.,2002;Wang et al.,2004);在小鼠肾间质细胞中,vhl蛋白缺失可导致肾脏皮质和髓质中EPO大量表达(Broeker et al.,2020)。在人类透明细胞肾细胞癌变细胞中,vhl蛋白失活后HIF激活葡萄糖转运蛋白(Glucose transporter type 1,GLUT1)调节葡萄糖摄取,并调控多种糖酵解酶和乳酸脱氢酶的表达,以增加糖酵解通量(Semenza et al.,1994;Chakraborty,2020)。在斑马鱼(Danio rerio)(van Rooijen et al.,2009)、瓦氏黄颡鱼(Pelteobagrus vachelli)(Zhang et al.,2017)和暗纹东方鲀(Takifugu fasciatus)(Li et al.,2019)等水产物种中进行低氧胁迫处理后,均可促使其vhl基因mRNA表达水平显著上升,表明vhl基因在鱼类低氧胁迫转录响应过程中发挥重要作用,但其对低氧胁迫的響应机制尚未明确。【本研究切入点】多鳞鱚(Sillago sihama)又名沙锥鱼,隶属于鲈形目(Perciformes)鱚科(Sillaginidae)鱚属(Sillago),主要分布于印度洋—西太平洋热带浅海,是一种广温、广盐性鱼类(薛泰强,2010;黄洋等,2013)。多鳞鱚肉质鲜美,营养价值高,深受消费者喜爱,是我国重要的经济鱼类之一(杜涛和黄洋,2009)。近年来由于过度捕捞,导致多鳞鱚种群数量锐减,产量降低,而急需通过人工养殖增加产量以满足市场需求。但多鳞鱚耐低氧能力差(窒息点约1.0 mg/L)、应激反应强(杜涛等,2009),难以实现大规模工厂化养殖,严重制约其养殖业发展。因此,亟待开展多鳞鱚vhl基因的相关研究以明确其低氧响应分子机制。【拟解决的关键问题】克隆多鳞鱚vhl基因cDNA全长,并分析vhl基因在多鳞鱚主要组织及低氧胁迫后鳃组织和心脏中的表达情况,为揭示多鳞鱚应对低氧的适应机制提供研究资料,也为今后开展多鳞鱚新品种选育提供候选基因。
1 材料与方法
1. 1 试验材料
供试多鳞鱚由广东海洋大学东海岛海洋生物研究基地提供。Prime Script RT reagent Kit with gDNA Eraser购自TaKaRa公司,Power Green qPCR Mix(with ROX)购自广州东盛生物科技有限公司。
1. 2 低氧胁迫处理
对多鳞鱚群体进行低氧胁迫预试验,发现同一群体中不同多鳞鱚个体间耐低氧能力存在一定差异,平均窒息点约1.0 mg/L,出现大量浮头现象时的溶解氧(DO)约1.3 mg/L。因此,正式试验的低氧胁迫DO设为1.3 mg/L。将480尾体型相近的1龄多鳞鱚随机分配到4个200 L水缸中(编号分别为1、2、3和4),每缸120尾,暂养1周后进行低氧胁迫处理。通过调节氮气和空气的充气速度而控制养殖水体DO水平,实时监测DO以符合试验需求。试验开始前测得每缸水体DO为8.0 mg/L,分别向1、2和3号缸水体持续通入氮气使其DO降至1.3 mg/L,低氧处理(1.3 mg/L)1 h后,取1号缸10尾浮头的多鳞鱚个体作为低氧胁迫1 h组(Hypoxia-1-h);低氧处理(1.3 mg/L)6 h后,取2号缸10尾正常活动的多鳞鱚个体作为低氧胁迫6 h组(Hypoxia-6-h);低氧处理6 h后再通入氧气使3号缸水体DO恢复至8.0 mg/L维持4 h后,取3号缸10尾正常活动的多鳞鱚个体作为恢复氧气组(Reoxygen-4-h);4号缸持续通入空气维持水体DO在8.0 mg/L,取出4号缸10尾正常活动的多鳞鱚个体作为对照组(CK)。采样前对鱼进行麻醉,采集对照组多鳞鱚的鳃组织、心脏、肝脏、肌肉、脑组织、卵巢和精巢等7个组织,以及Hypoxia-1-h组、Hypoxia-6-h组和Reoxygen-4-h组多鳞鱚的心脏和鳃组织,每组均采集3尾。采集样品经液氮速冻后转移到-80 ℃冰箱保存备用。
1. 3 总RNA提取及cDNA合成
通过TRIzol法提取总RNA,以1.0%琼脂糖凝胶电泳检测RNA完整性;参照Prime Script RT reagent Kit with gDNA Eraser说明将总RNA反转录合成cDNA,-20 ℃保存备用。
1. 4 目的基因克隆
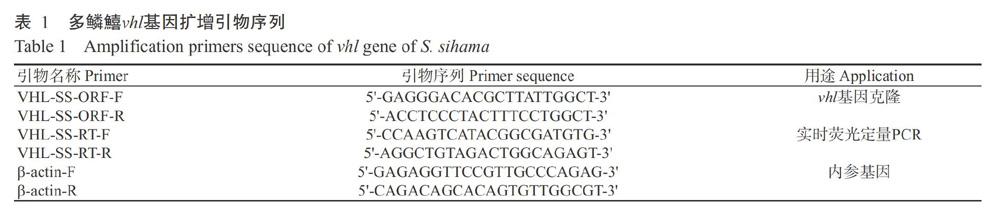
根据多鳞鱚转录组数据获取vhl基因序列(Tian et al.,2019),采用Primer Premier 6.0设计扩增引物(表1)。以对照组多鳞鱚的心脏、肝脏、脑组织和鳃组织混合cDNA为模板进行PCR扩增,反应体系25.0 μL:2×PCR Mix 12.5 μL,VHL-SS-ORF-F 1.0 μL,VHL-SS-ORF-R 1.0 μL,cDNA模板2.0 μL,双蒸水8.5 μL。扩增程序:94 ℃预变性3 min;94 ℃ 1 min,58 ℃ 30 s,72 ℃ 1 min,进行30个循环;72 ℃延伸10 min。PCR产物以1.0%琼脂糖凝胶电泳进行检测,凝胶成像系统检测扩增结果,并挑选单一明亮的扩增条带送至生工生物工程(上海)股份有限公司测序。
1. 5 生物信息学分析
使用EditSeq查找多鳞鱚vhl基因开放阅读框(ORF)并翻译成氨基酸序列,采用SMART(http://smart.embl-heidelberg.de/)预测多鳞鱚vhl蛋白结构域,以Signal IP 4.1(http://www.cbs.dtu.dk/services/SignalP/)预测多鳞鱚vhl蛋白前体信号肽,利用ClustalX比对多鳞鱚vhl氨基酸序列与其他脊椎动物的vhl氨基酸序列,并以MAGA 10.0中的邻接法(Neighbor-joining,NJ)构建系统发育进化树。
1. 6 多鳞鱚vhl基因组织半定量分析
以多鳞鱚的心脏、鳃组织、肝脏、脑组织、精巢、卵巢和肌肉共7个组织cDNA为模板,以VHL-SS-RT-F和VHL-SS-RT-R(表1)为引物、β-actin基因为内参基因,进行半定量PCR扩增。PCR产物以1.5%琼脂糖凝胶电泳进行检测,凝胶成像系统检测扩增结果,并根据条带亮度判断多鳞鱚vhl基因表达量。
1. 7 低氧胁迫后多鳞鱚vhl基因表达定量分析
将不同低氧胁迫组及CK组的多鳞鱚心脏和鳃组织cDNA进行5倍稀释,以VHL-SS-RT-F和VHL-SS-RT-R(表1)為引物、β-actin基因为内参基因,使用LightCycler 96实时荧光定量分析仪(Roche,USA)进行定量分析。反应体系20.0 μL:Power Green qPCR Mix(with ROX)10.0 μL,上、下游引物各0.8 μL,cDNA模板2.0 μL,双蒸水6.4 μL。扩增程序:95 ℃预变性10 min;95 ℃ 10 s,60 ℃ 30 s,72 ℃ 15 s,进行40个循环。每个样品设3个生物学重复和3个技术重复。以2-ΔΔCt法计算目的基因相对表达量,并通过SPSS 17.0进行单因素方差分析(One-way ANOVA)和Duncan?s多重比较。
2 结果与分析
2. 1 多鳞鱚vhl基因cDNA序列克隆及其序列分析结果
克隆获得的多鳞鱚vhl基因cDNA序列全长727 bp(GenBank登录号MN013394),包括120 bp的5'端非编码区(5'-TUR)、106 bp的3'端非编码区(3'-TUR)和504 bp的ORF,共编码167个氨基酸残基(图1)。多鳞鱚vhl基因编码蛋白主要有2个结构域,即Pfam:VHL(第12~93位氨基酸)和Pfam:VHL_C(第5~22、31~39和98~153位氨基酸),还包括ParB结构域(第100~162位氨基酸)和SOCS_box结构域(第104~140位氨基酸)。据Signal IP 4.1预测结果显示,多鳞鱚vhl蛋白无前体信号肽。
2. 2 多鳞鱚vhl氨基酸序列比对分析结果
vhl氨基酸序列BLASTp比对分析结果显示,多鳞鱚vhl氨基酸序列与盲曹鱼(Lates calcarifer)vhl氨基酸序列的同源性最高(81%),与高体鰤(Seriola dumerili)、攀鲈(Anabas testudineus)、尼罗罗非鱼(Oreochromis niloticus)的vhl氨基酸序列同源性较高(73%~78%),与爬行类、两栖类和哺乳类等高级脊椎动物的同源性为52%~54%(表2和图2)。从基于vhl氨基酸序列同源性构建的系统发育进化树(图3)也可看出,所有鱼类聚为一支,而爬行类、两栖类和哺乳类聚为一支。其中,多鳞鱚与鲈形目的高体鰤、盲曹鱼、尼罗罗非鱼和攀鲈先聚为一小分支,且与攀鲈的亲缘关系最近,与尼罗罗非鱼、攀鲈和高体鰤的亲缘关系次之,与斑点雀鳝(Lepisosteus oculatus)、斑马鱼(Danio rerio)和虹鳟(Oncorhynchus mykiss)等鱼类的亲缘关系相对较远。
2. 3 多鳞鱚vhl基因在不同组织中的表达情况
利用半定量PCR扩增检测多鳞鱚vhl基因在鳃组织、心脏、脑组织、肝脏、卵巢、精巢和肌肉等7个组织中的表达情况,结果(图4)显示,多鳞鱚vhl基因在不同组织中均有表达,其中以鳃组织、卵巢和精巢的表达量较高,其次是心脏和肝脏,在肌肉和脑组织的表达量较低。
2. 4 低氧胁迫对多鳞鱚vhl基因在鳃组织和心脏中表达的影响
实时荧光定量PCR检测结果(图5)显示,与CK组相比,Hypoxia-1-h组、Hypoxia-6-h组和Reoxygen-4-h组多鳞鱚vhl基因在鳃组织中的相对表达量显著上调(P<0.05,下同),但各低氧胁迫处理组间的vhl基因相对表达量无显著差异(P>0.05)。在心脏中,Hypoxia-1-h组和Hypoxia-6-h组多鳞鱚vhl基因的相对表达量较CK组均显著上调;Reoxygen-4-h组多鳞鱚vhl基因的相对表达量虽然较Hypoxia-6-h组显著下调,但仍显著高于CK组水平。
3 讨论
HIF介导的低氧信号通路是细胞产生低氧应答的核心信号通路之一(李福祥等,2004;Dunwoodie,2009;肖武汉,2014),而vhl在低氧信号通路中发挥重要作用。目前,关于vhl基因的研究集中在哺乳类动物中,针对鱼类的研究较少。本研究成功克隆获得多鳞鱚vhl基因cDNA序列全长727 bp,包括120 bp的5'-TUR、106 bp的3'-TUR和504 bp的ORF,共编码167个氨基酸残基。多鳞鱚vhl基因编码蛋白有4个结构域,分别是Pfam:VHL(第12~93位氨基酸)、Pfam:VHL_C(第5~22、31~39和98~153位氨基酸)、ParB(第100~162位氨基酸)和SOCS_box(第104~140位氨基酸)。其中,Pfam:VHL是由β-折叠组成的β-结构域(第12~93位氨基酸)(Stebbins et al.,1999);Pfam:VHL_C是由2个连接体(第5~22和31~39位氨基酸)和1个极性接口(第98~153位氨基酸)组成的α-螺旋结构域(Stebbins et al.,1999);在哺乳动物Sulfiredoxin-1蛋白中含有ParB结构域,而该蛋白有助于抗氧化应激(Chang et al.,2004;Figge et al.,2010);SOCS_box结构域是特定底物与蛋白结合的位点(Bullock et al.,2007),如E3泛素蛋白连接酶。上述结构域功能进一步证实vhl在低氧信号通路中发挥重要作用。多鳞鱚与其他脊椎动物的vhl氨基酸序列同源性比对分析结果表明,多鳞鱚与盲曹鱼的同源性最高,为81%,与鲈形目其他鱼类的同源性为73%~78%,与其他硬骨鱼类的同源性为60%~67%,与爬行类、两栖类和哺乳类的同源性较低(52%~54%)。系统发育进化分析结果也显示,多鳞鱚与鲈形目鱼类聚为一支,即vhl的系统发育进化关系与分类学进化关系一致。
已有研究发现,vhl基因在瓦氏黄颡鱼(Zhang et al.,2017)和暗纹东方鲀(Li et al.,2019)各组织中均有表达分布。本研究同样发现,vhl基因在多鳞鱚主要组织中均有表达。其中,多鳞鱚vhl基因在鳃组织、卵巢和精巢中高度表达,在肝脏和心脏中的表达量次之,而在肌肉和脑组织中的表达量较低。但vhl基因在暗纹东方鲀和瓦氏黄颡鱼中均以肝脏和心脏的表达量较高,而在脑组织和鳃组织的表达量相对较低(Zhang et al.,2017;Li et al.,2019)。vhl基因在多鳞鱚各组织中的表达量与暗纹东方鲀和瓦氏黄颡鱼存在差异,说明vhl基因表达可能具有物种特异性。
本研究对比分析低氧胁迫前后多鳞鱚vhl基因在鳃组织和心脏中的表达变化,结果显示,低氧胁迫1 h后多鳞鱚vhl基因在鳃组织和心脏中的表达量均显著上调,但在瓦氏黄颡鱼和暗纹东方鲀脑组织及肝脏中的表达量无显著差异(Zhang et al.,2017;Li et al.,2019),表明不同鱼类对低氧产生应答的时间也不同;低氧胁迫6 h后多鳞鱚vhl基因在心脏中的表达量持续显著上升,在瓦氏黄颡鱼和暗纹东方鲀肝脏及脑组织中也有相似的表达规律(Zhang et al.,2017;Li et al.,2019),表明vhl基因在不同鱼类低氧胁迫转录响应中均发挥重要作用。低氧胁迫6 h后,多鳞鱚vhl基因在鳃组织中的表达量较低氧胁迫1 h时无显著差异,在暗纹东方鲀肝脏中有相同的表达模式,但暗纹东方鲀在低氧胁迫8 h后vhl基因在肝脏中的表达极显著上升(Li et al.,2019),可能是vhl基因在不同組织间对低氧胁迫产生应答的时间不同。恢复氧气4 h后,多鳞鱚vhl基因在鳃组织中的表达量与低氧胁迫时无显著变化,在心脏中的表达量较低氧胁迫6 h时显著下调,但仍显著高于正常水平。Zhang等(2017)在瓦氏黄颡鱼肝脏中观察到与多鳞鱚鳃组织相同的表达模式,Li等(2019)则在暗纹东方鲀脑组织和肝脏中观察到与多鳞鱚心脏相同的表达模式,可能与鱼体在低氧胁迫再恢复氧气后不同组织所处应激状态或受损程度不一致有关。
鳃组织作为鱼类感受低氧胁迫最直接的器官,会以最快速度减轻低氧胁迫对机体的影响;而心脏作为鱼类生命活动的重要器官,是在保证组织损伤程度最小的前提下对低氧进行响应。在本研究中,低氧胁迫下多鳞鱚vhl基因在鳃组织和心脏中的应答速度及表达稳定性存在一定差异。在鳃组织中,多鳞鱚vhl基因的表达应答速度快,各低氧胁迫处理组与CK组间存在显著差异,但各低氧胁迫处理组间的表达量稳定,无显著变化。鳃组织在急性低氧胁迫环境下表现出强烈的结构改变,包括鳃小片肿胀、抬升及细胞排列发生紊乱等(狄治朝等,2018);低氧胁迫再恢复氧气后,鳃组织结构虽然能得到一定程度的恢复(杨明等,2019),但恢复程度与能力可能存在物种差异。多鳞鱚在低氧胁迫下其鳃组织结构可能已发生较严重损伤,即使恢复氧气相关功能基因感知DO的响应能力仍较弱,表现为各低氧胁迫处理组间vhl基因表达量较稳定,无显著差异。在心脏中,多鳞鱚vhl基因的表达应答速度较鳃组织慢,各低氧胁迫处理组与CK组间也存在显著差异。心脏应答速度慢,可能是因为鱼类的心脏耐低氧能力较强,能适应极端的低氧条件(Marques et al.,2008)。随着低氧胁迫时间的延长,vhl基因在心脏中的表达量持续升高,但恢复氧气后其表达量下降,可能是心脏为提高机体携氧能力而做出的应激反应。综上所述,vhl基因在鱼类低氧胁迫转录应答中发挥重要作用,且在不同物种不同组织间的表达存在差异,即vhl基因功能可能具有组织特异性和物种特异性,但具体原理有待进一步探究。
4 结论
多鳞鱚vhl基因编码蛋白存在Pfam:VHL、Pfam:VHL_C、ParB和SOCS_box等4个结构域,且低氧胁迫前后其表达量存在显著差异,即多鳞鱚vhl基因在低氧应答信号通路中扮演着重要角色。
參考文献:
杜涛,黄洋. 2009. 多鳞鱚生物学特性及室内养殖试验[J]. 水产养殖,(3):1-3. [Du T,Huang Y. 2009. Biological cha-racteristics and indoor multiplication experiment of Sillago sihama Forskál[J]. Journal of Aquaculture,(3):1-3.]
杜涛,黄洋,曹剑香. 2009. 多鳞鱚采捕暂养的初步研究[J]. 养殖与饲料,(10):15-17. [Du T,Huang Y,Cao J X. 2009. Preliminary study on harvesting and temporary maintenance of Sillago sihama Forskál[J]. Animals Breeding and Feed,(10):15-17.]
狄治朝,周涛,许强华. 2018. 低氧胁迫与常氧条件下斑马鱼鳃中热休克蛋白基因家族的表达差异比较[J]. 大连海洋大学学报,33(6):690-695. [Di Z C,Zhou T,Xu Q H. 2018. Expression comparisons of heat shock proteins gene in gills of zebrafish exposed to both normoxia and hypoxia[J]. Journal of Dalian Ocean University,33(6):690-695.]
黄洋,杜涛,黄海立. 2013. 多鳞鱚Sillago sihama Forskál人工繁殖研究[J]. 广东海洋大学学报,33(1):15-21. [Huang Y,Du T,Huang H L. 2013. A study on artificial bree-ding of Sillago sihama Forskál[J]. Journal of Guangdong Ocean University,33(1):15-21.]
李福祥,全燕,夏前明. 2004. 低氧诱导因子及其低氧调节机制[J]. 中国临床康复,8(30):6744-6746. [Li F X,Quan Y,Xia Q M. 2004. Hypoxia inducible factor and its hypoxia regulating mechanism[J]. Chinese Journal of Clinical Rehabilitation,8(30):6744-6746.]
武晓会,刘洋,狄治朝,赵文静,王丛丛,许强华. 2018. 低氧对斑马鱼细胞存活能力的影响[J]. 南方农业学报,49(8):1641-1647. [Wu X H,Liu Y,Di Z C,Zhao W J,Wang C C,Xu Q H. 2018. Effects of hypoxia on cell survival ability of zebra fish[J]. Journal of Southern Agriculture,49(8):1641-1647.]
肖武汉. 2014. 低氧信号传导途径与鱼类低氧适应[J]. 中国科学:生命科学,44(12):1227-1235. [Xiao W H. 2014. Hypoxia signaling pathway and hypoxic adaptation of fishes[J]. Science China-Life Sceences,44(12):1227-1235.]
薛泰强. 2010. 鱚科几种鱼类的形态学及遗传学研究[D]. 青岛:中国海洋大学. [Xue T Q. 2010. Study on morphology and genetics of some Sillaginidae species[D]. Qingdao:Ocean University of China.]
杨明,孙盛明,傅洪拓,乔慧,张文宜,龚永生,蒋速飞,熊贻伟,许蕾,赵才源,王亚冰,胡宇宁. 2019. 低氧和复氧对日本沼虾抗氧化酶活力及组织结构的影响[J]. 中国水产科学,26(3):493-503. [Yang M,Sun S M,Fu H T,Qiao H,Zhang W Y,Gong Y S,Jiang S F,Xiong Y W,Xu L,Zhao C Y,Wang Y B,Hu Y N. 2019. Hypoxia and reoxygenation on antioxidant enzyme activities and histological structure of Macrobrachium nipponense[J]. Journal of Fishery Sciences of China,26(3):493-503.]
Ankoma-Sey V,Wang Y,Dai Z. 2000. Hypoxic stimulation of vascular endothelial growth factor expression in activated rat hepatic stellate cells[J]. Hepatology,31(1):141-148.
Broeker K A E,Fuchs M A A,Schrankl J,Kurt B,Nolan K A,Wenger R H,Kramann R,Wagner C,Kurtz A. 2020. Different subpopulations of kidney interstitial cells produce erythropoietin and factors supporting tissue oxyge-nation in response to hypoxia in vivo[J]. Kidney International. doi: 10.1016/j.kint.2020.04.040.
Bullock A N,Rodriguez M C,Debreczeni J ?,Zhou S Y,Knapp S. 2007. Structure of the SOCS4-ElonginB/C complex reveals a distinct SOCS box interface and the mole-cular basis for SOCS-dependent EGFR degradation[J]. Structure(Cambridge),15(11):1493-1504.
Chakraborty A A. 2020. Coalescing lessons from oxygen sen-sing,tumor metabolism,and epigenetics to target VHL loss in kidney cancer[J]. Seminars in Cancer Biology. doi: 10.1016/j.semcancer.2020.03.012.
Chang T S,Jeong W,Woo H A,Lee S M,Park S,Rhee S G. 2004. Characterization of mammalian sulfiredoxin and its reactivation of hyperoxidized peroxiredoxin through reduction of cysteine sulfinic acid in the active site to cysteine[J]. The Journal of Biological Chemistry,279(49):50994-51001.
Corpechot C,Barbu V,Wendum D,Kinnman N,Rey C,Poupon R,Housset C,Rosmorduc O. 2002. Hypoxia-induced VEGF and collagen I expressions are associated with angiogenesis and fibrogenesis in experimental cirrhosis[J]. Hepatology,35(5):1010-1012.
Dunwoodie S L. 2009. The role of hypoxia in development of the mammalian embryo[J]. Developmental Cell,17(6):755-773.
Figge R M,Easter J,Gober J W. 2010. Productive interaction between the chromosome partitioning proteins,ParA and ParB,is required for the progression of the cell cycle in Caulobacter crescentus[J]. Molecular Microbiology,47(5):1225-1237.
Kaelin W G. 2007. The von Hippel-Lindau tumor suppressor protein and clear cell renal carcinoma[J]. Clinical Cancer Research,13(2):680s-684s.
Li X R,Wang T,Yin S W,Zhang G S,Cao Q Q,Wen X,Zhang H Y,Wang D,Zhu W X. 2019. The improved energy metabolism and blood oxygen-carrying capacity for pufferfish,Takifugu fasciatus,against acute hypoxia under the regulation of oxygen sensors[J]. Fish Physiology and Biochemistry,45(1):323-340.
Marques I J,Leito J T D,Spaink H P,Testerink J,Jaspers R T,Witte F,Berg S V,Bagowski C P. 2008. Transcriptome analysis of the response to chronic constant hypo-xia in zebrafish hearts[J]. Journal of Comparative Physio-logy,178(1):77-92.
Pollock M S,Clarke L M J,Dubé M G. 2007. The effects of hypoxia on fishes:From ecological relevance to physiological effects[J]. Environmental Reviews,15:1-14.
Roberts J J,Brandt S B,Fanslow D,Ludsin S A,Pothoven S A,Scavia D,H??k T O. 2011. Effects of hypoxia on consumption,growth,and RNA:DNA ratios of young yellow perch[J]. Transactions of the American Fisheries Society,140(6):1574-1586.
Semenza G L,Roth P H,Fang H M,Wang G L. 1994. Transcriptional regulation of genes encoding glycolytic enzymes by hypoxia-inducible factor 1[J]. The Journal of Biological Chemistry,269(38):23757-23763.
Stebbins C E,Kaelin Jr W G,Pavletich N P. 1999. Structure of the VHL-ElonginC-ElonginB complex:Implications for VHL tumor suppressor function[J]. Science,284(5413):455-461.
Tian C X,Li Z Y,Dong Z D,Huang Y,Du T,Chen H P,Jiang D N,Deng S P,Zhang Y L,Wanida S,Shi H J,Wu T L,Zhu C H,Li G L. 2019. Transcriptome analysis of male and female mature gonads of silver sillago(Sillago sihama)[J]. Genes,10(2):129.
van Rooijen E,Voest E E,Logister I,Korving J,Schwerte T,Schulte-Merker S,Giles R H,van Eeden F J. 2009. Zebrafish mutants in the von Hippel-Lindau tumor suppressor display a hypoxic response and recapitulate key aspects of Chuvash polycythemia[J]. Blood,113(25):6449-6460.
Wang Y Q,Luk J M,Ikeda K,Man K,Chu A C,Kaneda K,Fan S T. 2004. Regulatory role of vHL/HIF-1α in hypoxia-induced VEGF production in hepatic stellate cells[J]. Biochemical and Biophysical Research Communications,317(2):358-362.
Zhang G S,Zhao C,Wang Q T,Gu Y C,Li Z C,Tao P F,Chen J W,Yin S W. 2017. Identification of HIF-1 signa-ling pathway in Pelteobagrus vachelli using RNA-Seq:Effects of acute hypoxia and reoxygenation on oxygen sensors,respiratory metabolism,and hematology indices[J]. Journal of Comparative Physiology,187(7):931-943.
(責任编辑 兰宗宝)
