
低温胁迫下贵州云雾贡茶生长调节剂的变化
2020-07-14 08:33王莹李岩王姝向准赵德刚
湖北农业科学 2020年8期
王莹 李岩 王姝 向准 赵德刚
摘要:以贵州云雾贡茶[ Camellia sineWis(L.)Kuntze var.niaowangen.sis Q.H.Chen] -年生幼苗茶树为试验材料,用酶联免疫法(ELISA)测定不同低温胁迫下茶树叶片生长调节剂含量的变化,以探讨低温胁迫下茶树生长调节剂变化规律。结果显示,l、4。C处理云雾贡茶IAA、GA含量大部分显著低于对照,ABA含量、ABA/GA明显高于对照;10℃处理ABA、IAA、GA含量大部分显著高于对照,ABA/GA与对照无明显变化。随胁迫时间延长,1、4℃处理ABA、IAA含量随时间变化呈先上升后下降的趋势,GA含量呈下降趋势,ABA/GA呈上升趋势;10℃处理ABA和GA含量呈先上升后下降的趋势,IAA含量呈下降趋势。低温奈件下,云雾贡茶能提高ABA类生长抑制物质,降低IAA、GA等生长促进类物质,通过调节植物体内生长调节剂平衡来应对低温胁迫。
关键词:云雾贡茶[ Camellia sinensis(L)Kuntze var.niaowangensis Q.H.Chen];低温胁迫;生长调节剂
中图分类号:S571.1; Q946.885
文献标识码:A
文章编号:0439-8114( 2020) 08-0099-04
D01:10.1408 8/j.cnki.issn0439- 8114.2020.08.022
云雾贡茶[Camellia sinensis (L.) Kuntze var.niaowangensisQ.H.Chen]良种分布在贵州省贵定县南部云雾山地区,云雾山区域独特的生态环境孕育了云雾贡茶独特茶香嫩栗香,富含有机锗、茶多糖。云雾贡茶栽培种植历史悠久,早在唐朝就被茶圣陆羽收录记载,元泰定二年(公元1325年)开始作为朝廷贡茶,至明清更成为皇家珍品,乾隆时列为全国八大名茶之一。近年来,用云雾贡茶制作的贵定雪芽连获国际金奖,成为国际名茶,是国内目前仅存贡茶碑记载的名优茶种[1]。
低温是影响茶树生长和繁殖的重要因素,冬季的极端低温和初春的霜冻或倒春寒常常造成茶树的低温伤害,制约茶叶产业发展[2,3]。目前,针对植物抗寒性,大多集中于细胞膜透性、保护酶系统及渗透性物质含量变化等方面的研究[4-6],生长调节剂与抗寒性关系研究较少[7]。当植物遭受低温等逆境胁迫时,植物生长调节剂如ABA、GA等被认为是植物对抗寒基因表达的启动因子[8],对增强植株抗寒性起重要作用。本研究测定分析了贵州云雾贡茶在不同低温胁迫下叶片中生长调节剂的含量,以探讨低温胁迫下云雾贡茶生长调节剂的变化规律,从而为深入研究云雾贡茶耐寒机理与其生长调节剂的关系提供依据。
1 材料与方法
1.1 材料
供试材料为云雾贡茶一年生扦插苗,材料统一栽植于塑料盆中,每盆1株。在日光温室内自然光照下进行正常管理,当植株长至株高70-80 cm时,选取健康状况良好、长势一致的植株进行试验处理[7]。
1.2试验设计
将供试幼苗分为4组,每组8盆,于试验第1天转入RX2-3008型人工气候箱中,人工气候箱的温度分别设为1、4、10、25℃,以25℃处理为对照(CK),光周期昼/夜为16 h/8 h,光照度为12 000 1x,相对湿度为50%。于胁迫处理的1、3、5、7、9d选取功能叶片测定生长调节剂的含量。
1.3 生长调节剂的测定
生长调节剂脱落酸(ABA)、生长素(IAA)、赤霉素(CA)含量采用酶联免疫吸附测定法(EIISA)测定,试剂盒采购于中国农业大学植物生理与生物化学国家重点实验室。
1.4 数据分析
数据分析采用Excel和SPSS软件,采用Duncan新复极差法对处理平均数间的差异显著性进行检验。
2 结果与分析
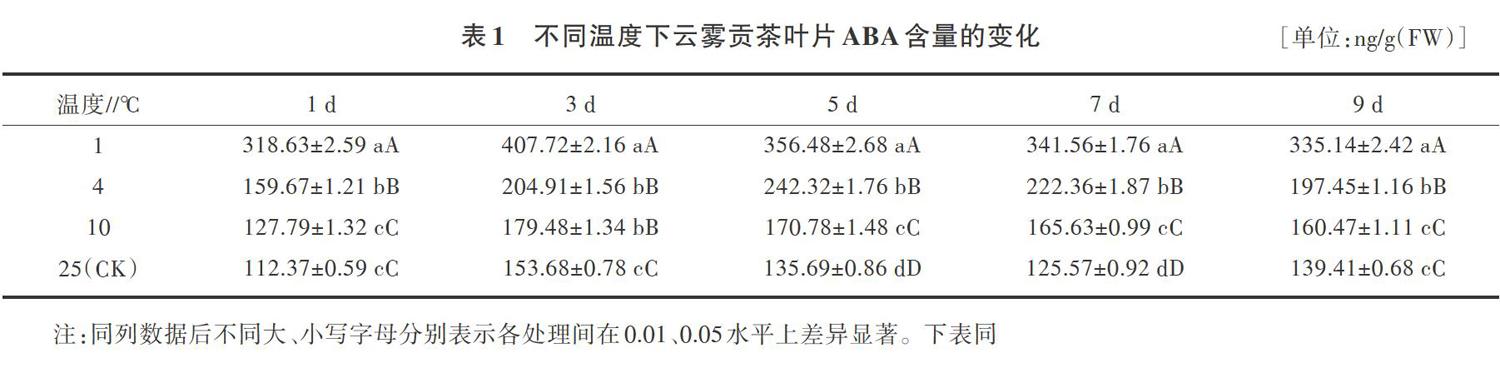
2.1 低温胁迫下云雾贡茶叶片ABA含量的变化
逆境下,生长调节剂ABA含量显著增加,以调节植物对胁迫环境的适应[9]。结果(表1)表明,在同一胁迫时间内,随着胁迫温度的降低,云雾贡茶ABA含量呈上升趋势;随着胁迫时间的延长,相同低温处理下,云雾贡茶的ABA含量均呈先上升后下降的趋势,1、4、10℃处理分别在3、5、7d含量达到最高,与对照相比分别提高了165.3%、78.6%、16.8%。1、4、10℃處理与对照相比差异极显著。
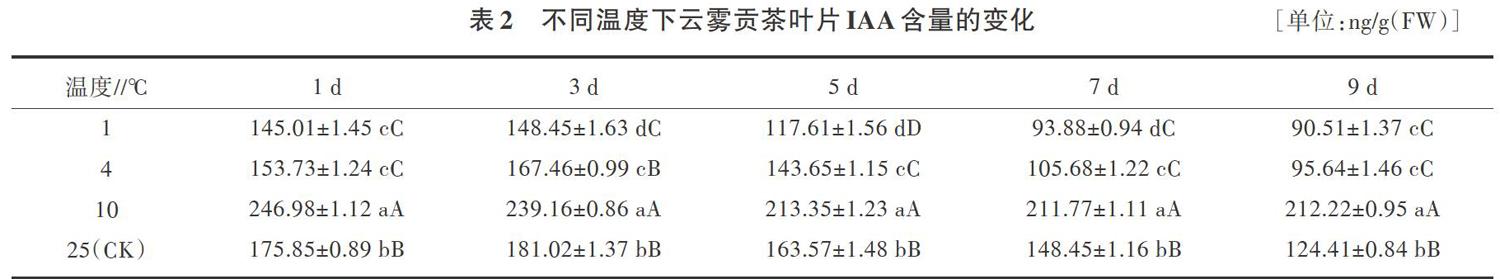
2.2 低温胁迫下云雾贡茶叶片IAA含量的变化
IAA是植物体最普遍的生长素类物质,能促进细胞的分裂、伸长和分化[7]。结果(表2)表明,在不同胁迫时间下,除3 d外,1、4℃的低温处理云雾贡茶的IAA含量与对照相比均有大幅度下降,差异达到极显著水平;不同胁迫时间下,10℃的低温处理云雾贡茶的IAA含量与对照相比均有大幅度上升,差异达到极显著水平。同一胁迫温度下,随着胁迫时间的延长,IAA含量大多数呈先上升后下降的趋势,1、4℃胁迫处理9d时,IAA含量比对照分别降低了27.25%、23.13%。
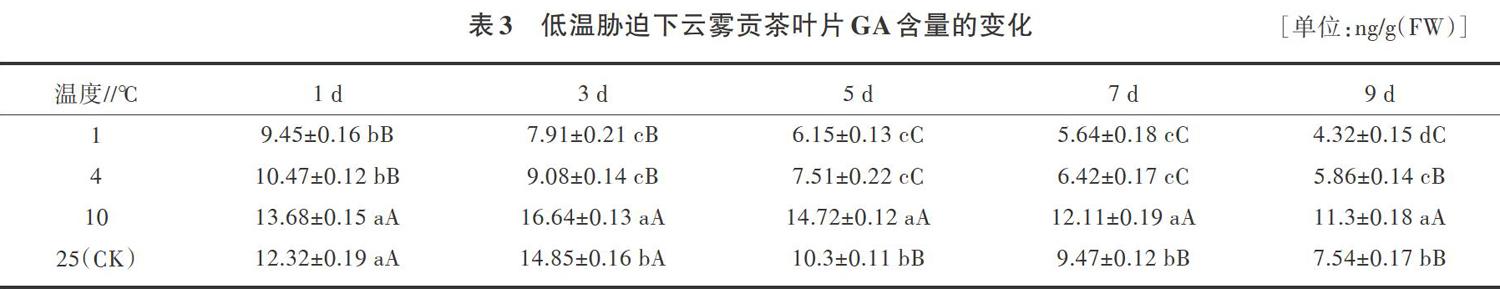
2.3 低温胁迫下云雾贡茶叶片GA含量的变化
从表3可以看出,随着胁迫时间的延长,1、4℃处理下云雾贡茶GA含量呈下降的趋势,10、25℃处理下,云雾贡茶GA含量呈先上升后下降的趋势,10℃处理均高于25℃对照处理。1、4℃处理与对照相比,GA含量差异达到极显著水平(胁迫9d除外),胁迫9d时,GA含量比对照分别降低了42.71%、22.29%。CA含量的下降能抑制植物生长,提高抗寒性,这表明1、4℃处理云雾贡茶能通过改变体内GA的含量来适应低温的变化。
2.4 低温胁迫下云雾贡茶叶片ABA/GA的变化
植物抗寒力与植物生长调节剂间的平衡有关,ABA与赤霉素类(GAs)间的平衡被看作是植物应对胁迫时的重要调节因子[10],一般情况下,植物组织中高比率的ABA/GAs意味着较高的抵抗寒冷的能力[11]。
由图1可知,同一胁迫时间下,随着胁迫温度的降低,云雾贡茶各处理的ABA/GA明显升高;随胁迫时间延长,1、4℃处理ABA/GA呈上升趋势,且1℃处理ABA/GA上升较快,4℃处理上升趋势缓慢;10℃处理与对照相比ABA/GA无明显变化。
3 小结与讨论
植物生长调节剂在生物信号间存在相互作用,包括生物合成、代谢与运输、信号转导途径等方面的相互作用[12-15]。近年来,ABA已经被认为是一种胁迫生长调节剂,在植物抗寒力的调控以及对维持细胞膜结构功能方面起着重要作用;植物受到生物和非生物胁迫后,内源ABA含量增加,并诱导多种抗逆基因的表达,从而增强其抗逆性;ABA含量的增加对于植物抗性的提高有明显作用[16-20]。同一胁迫时间内,随着胁迫温度的降低,云雾贡茶的ABA含量呈上升趋势,这可能是茶树在对抗低温时的一种保护性反应。随胁迫时间延长,各低温处理下ABA含量均呈先上升后下降的趋势,可能是因为植物的ABA含量持续性积累到一定程度后不能再继续合成,到了胁迫后期植物的细胞膜、保护酶系统等已经受到了一定程度的伤害[11]。
生长素是最早发现的一类植物生长调节剂,它调控侧根形成、维管组织分化、顶端优势以及植物的向性反应等[20]。本试验中,随着胁迫时间的延长,IAA含量大多数呈先上升后下降的趋势。在1、4℃的低温处理下云雾贡茶的IAA含量与对照相比均下降;10℃的低温处理下,IAA含量与对照相比上升。这与小麦[21]、珙桐[22]IAA含量对低温的反应是一致的,但也有研究表明,低温胁迫IAA含量呈上升趋势[23],这可能与试验材料与处理条件的差异、外部环境条件的影响等有关系。
最早被认为与抗寒力有关的植物生长调节剂是赤霉素类,研究表明抗寒性强的植物体内赤霉素的含量一般比抗寒性弱的植物低[24]。本研究结果表明,同一胁迫温度下,随着胁迫时间的延长,云雾贡茶的GA含量均有不同程度的下降;1、4℃处理均低于对照,10℃处理高于对照。这证明云雾贡茶能通过降低GA含量减缓植株生长速度,从而适应低温的胁迫。
Zhang等[25]引认为ABA/CA之间的平衡是提高植物抗寒性的重要因子之一。本研究中,同一胁迫时间下,云雾贡茶ABA/GA随胁迫温度降低而明显升高;随着胁迫时间的延长,在1、4℃处理呈上升趋势,变化幅度较大;10℃和对照变化不明显。Pocie-cha等[26]认为ABA/GA值的增加与植物抗寒性的增强呈密切正相关,但不同品种或同种植物不同生境下ABA、GA的贡献率不同,本研究中云雾贡茶ABA/GA变化主要与ABA的变化有关。
茶树生长调节剂虽然含量很低,但它们在调节茶树的生长、生理和新陈代谢等各方面起重要的调控作用[20]。在不同低温胁迫条件下,研究云雾贡茶生长调节剂的变化、相互关系及其对应基因的表達,有助于了解云雾贡茶对低温胁迫的响应机理,从而为挖掘茶树抗寒基因提供理论依据。
参考文献:
[1]王济红,陈谦海,林昌虎.云雾贡茶——贵州山茶属一新变种[J-种子,2011.30(1):65-66
[2]邹中伟,房婉萍,张定,等.低温胁迫下茶树基因表达的差异分析[J].茶叶科学,2008.28(4):249-254.
[3]朱全武,范凯,谢艳兰,等.植物低温胁迫响应miRNAs及其在茶树抗寒研究中的应用[J].茶叶科学,2013.33(3):212-220.
[4]何跃君,薛立,任向荣,等.低温胁迫对六种苗木生理特性的影响[J].生态学杂志,2008.27(4):524-531.
[5]申亚梅,马进,黄玉英,等.低温胁迫对5种园林常绿灌木膜透性的影响[J].浙江林业科技,2009.28(6):28-31.
[6]郁万文,曹福亮,汪贵斌.低温胁迫下银杏活性氧代谢与膜伤害的关系[J].东北林业大学学报,2010.38(7):46-48.
[7]张迎辉,李书平,荣俊冬,等.低温胁迫下福建山樱花、日本樱花内源激素的变化[J].福建林业科技,2013 .40(3):62-67.
[8] WEYERS J D B. PATERSON N W. Plant hormone and the controlof physiological processes EJl.New phytologist, 2002. 152 (3):375-407.
[9] CUSTA L,TRISCHUK R. WEISER C J.Plant cold acclimation:The role of ahscisic acid [Jl.Journal of plant growth regulation,2005.24(4):308-318.
[10] DAIE J, CAMPBELL W F.Response of tomato plants to stressfultemperatures increase in abscisic acid concentrations[J]. Plantphysiology, 1981 .67(1):26-29.
[11]张迎辉.低温胁迫下福建山樱花的生理响应与抗寒基因的表达[D].福州:福建农林大学,2014.
[12] DU H. WU N. CHANG Y. et al.Carotenoid deficiency impairsABA and IAA hiosvnthesisand differentiallv affects drought andcold tolerance in rice[J]. Plant molecular hiology, 2013, 83 (4):475-488.
[13] GOLENBERG E W. WEST N W. Hormonal interactions and generegulation can link monoecy and environmental plasticity to theevolution of dioecy in plants [ J] . American journal of botany . 2013 ,100( 6) :1022-1037.
[14] MIRANSARI M. SMITH D L. Plant hormones and seed germina-tion[J]. Environmental and experimental botany,2014,99: 1 10- 121.
[15] CHENG Y Q. LIU J F, YANG X D . et al. Construction of ethyleneregulatory network hased on the phytohormones related gene tran-scriptome profiling and prediction of transcription factor activitiesin soybean [J].Acta physiologiae plantarum. 2013. 35 (4) :1303 -1317.
[16] CAO F Y. YOSHIOKA K. DESVEAUX D. The roles of ABA inplant-pathogen interactions [ J ] . Journal of plant research . 201 1 ,124(4) :489-499.
[17] LEE S C , LUAN S. ABA signal transduction at the crossroad of bi-otic and abiotic stress responses [Jl. Plant cell and environment.2012.35( 1) : 53-60.
[18] NAKAMURA T. YAZAKI J. KISHIMOTO N. et al. Comparison oflong-term up-regulated genes during induction of freezing toler-ance by cold and ABA in bromegrass cell cultures revealed by mi-croarray analyses [Jl. Plant growth regulation, 2013. 71 (2) :113-136.
[19] DANQUAH A, ZELICOURT A D. COLCOMBET J, et al. The roleof ABA and MAPK signaling pathways in plant abiotic stress re-sponses[ J] . Plant biotechnology. 2014 . 32( 1 ) : 40-52.
[20]曾光辉,马青平,王伟东,等,自然低温对茶树内源激素含量的影响 [J].茶叶科学 , 2016.36( 1) : 85-91.
[21] KOSOVA K. PRASIL I T, VITDMV P. et al. Complex phytohor-mone responses during the cold acclimation of two wheat cultivarsdiffering in cold tolerance . winter Samanta and spring Sandra [J ] .Journal of plant physiology , 2012 . 169( 6) : 567-576.
[22] GUO B, STILES A R. LIU C Z. Changes in endogenous hormonesand oxidative burst as the biochemical basis for enhanced shoot or-ganogenesis in cold-treated Saussurea involucrata explants [ J ] . Ac-to physiologiae plantarum. 2013 . 35 ( 1 ) : 283-287.
[23]歐阳琳,洪亚辉,黄丽华,等,不同逆境胁迫信号对超级稻幼凿生理生化影响及植物激素变化的初步研究 [Jl.业现代化研究 , 2007.28( 1) : 104-106.
[24] 罗正荣,植物激素与抗寒力的关系 [Jl.植物生理学通讯 . 1989(3):1-5
[25 ] ZHANG F.WAN X Q.ZHANG H Q, et al. The effect of cold stresson endogenous hormones and CBFl homolog in four contrastingbamboo species [J]. Journal of forest research. 2012. 17 (1) :72-78.
[26] POCIECHA E . PLAIEK A. JANOWIAK F. et al. ABA level, pro- line and phenolic concentration. and PAL activity induced during cold acclimation in androgenic festulolium forms with contrastingresistance to frost and pink snow mould (Microdochitan nivale ) [ J ] .Physiological and molecular plant pathology, 2008. 73 (6) :126-132.
基金项目:贵州省高层次创新型人才培养“千层次”计划项目;山地植物资源保护与种质创新省部共建教育部重点实验室开放基金资助项目(MOELP-201703);贵州省科技基金项目[黔科合基础(2018)1044]
作者简介:王莹(1982-),女,黑龙江齐齐哈尔人,副研究员,在读博士研究生,主要从事植物分子生物学遗传育种方面的研究,(电话)18111862560(电子信箱)wangyinghouzi@163.com;通信作者,赵德刚(1961-),男,教授,博士生导师,主要从事植物基因T程研究, (电子信箱)dgzhao@gzu.edu.cn。
猜你喜欢
音乐天地(音乐创作版)(2022年6期)2022-10-14
作文小学中年级(2021年2期)2021-03-15
中国地市报人(2019年9期)2019-09-16
杂文月刊(选刊版)(2018年5期)2018-05-14
杂文选刊(2018年5期)2018-05-09
新疆农垦科技(2016年2期)2016-08-21
民族大家庭(2016年5期)2016-03-20
中国果菜(2016年9期)2016-03-01
中国马铃薯(2015年5期)2016-01-09
国学(2015年9期)2015-05-30
