
长江口北支水域甲壳类群落结构特征
2020-07-31 00:19赵蓬蓬高春霞陈锦辉戴黎斌王学昉
水产科学 2020年4期
赵蓬蓬,高春霞,2,3,4,陈锦辉,5,赵 静,2,3,4,戴黎斌,王学昉,2,3,4
( 1.上海海洋大学 海洋科学学院,上海 201306; 2.国家远洋渔业工程技术研究中心,上海 201306; 3.大洋渔业资源可持续开发教育部重点实验室,上海 201306; 4.农业农村部大洋渔业开发重点实验室,上海 201306; 5.上海市水生野生动植物保护研究中心,上海 202150 )
长江口生态环境复杂多样,温、盐度变化剧烈,具有丰富的营养盐类,是我国最大的淤泥质型河口生态系统[1]。因沿岸流、潮流、径流等影响,形成了其独特的地理环境[2],不仅渔产潜力极高,而且是鱼类洄游及虾蟹种类的索饵、产卵和育肥之地[3]。然而,近年来人类大型涉水工程和湿地围垦等因素导致长江口北支径流面积逐年减少,河床不断淤积且盐水倒灌南支现象使得该地域的群落结构和生物多样性呈现变动消长态势[4],造成生物群落结构发生重大变化[5]。与之相伴的是长江口海区陆源性物质增多[6],浮游生物过度繁殖,赤潮现象频繁发生,破坏了当地生态系统稳定性,严重影响了生态系统中甲壳类的群落结构[7]。
水生甲壳类经济、食用价值较高,不仅是重要的大型底栖动物,也是一种反映环境变迁的指示生物,其生物多样性特征和群落结构变化可有效预测河口生态环境变化[8]。目前,长江口大型底栖动物的研究大多集中在南支、湿地和口外海域[9-13],然而长江口北支属于口内海域,有关口内海域甲壳类生物多样性及群落的研究却少有报道[14]。为此,笔者基于2011─2014年渔业资源调查数据对长江口北支甲壳类资源的种类组成、群落结构多样性等进行分析,旨在为长江口生物多样性保护和生态环境管理提供理论支持。
1 材料与方法
1.1 数据来源
数据来源于2011年2月─2014年11月的长江口北支渔业资源底拖网调查,调查时间为每年的2月(冬季)、5月(春季)、8月(夏季)和11月(秋季),共4个航次,每个航次均设置5个采样点(图1)。采样方式为定点采样,调查船为沪崇渔1511号,其网具参数为:双囊底拖网,网口宽6 m,网高2 m,网纲长6 m,囊网网目20 mm。根据GPS进行站点定位,当调查船到达站位后,下拖网,以3 kn的航速拖曳,每个站点拖网30 min。所有渔获种类均按照《长江河口大型底栖无脊椎动物》进行鉴定[15],并在实验室进行种类鉴定与体质量、体长等常规生物学测定。
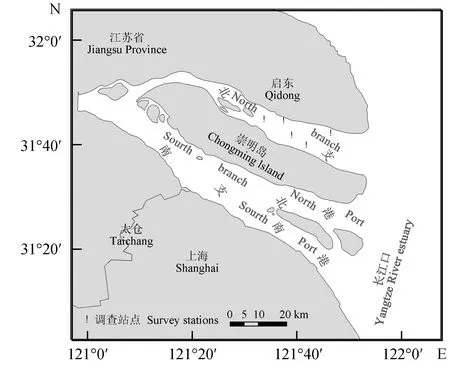
图1 长江口北支渔业资源调查站点分布
1.2 数据统计与分析
1.2.1 优势度
采用物种优势度(Y)对不同种类在群落中的重要性进行评价,当Y>0.02[16]时,即认为该物种为优势种,公式如下:
式中,N为各采样点所有物种个体总数,ni为第i种的个体总数,fi为该物种在各个采样点出现的频率。
1.2.2 物种多样性
采用Margalef丰富度指数(D)、Pielou均匀度指数(J)和Shannon-Wiener多样性指数(H′)对该水域甲壳类资源的物种多样性进行分析,公式
如下:
式中,N为总物种数,S为种类数,Pi为第i种占总尾数的比例。
1.2.3 群落结构分析
基于2011─2014年长江口北支甲壳类丰度数据构建群落的Bray-Curtis相似性系数矩阵,采用等级聚类分析和非度量多维标度排序方法对长江口北支甲壳类群落结构的相似程度进行分析[17-18],以此研究甲壳类群落结构组成的相似性;并依据群落更新指数、迁移指数和丰度—生物量比较曲线的变化情况[19-20],分析甲壳类群落结构的稳定性。以上所有数据处理均在ArcGIS 10.3和R 3.6.1软件中进行。
2 结 果
2.1 种类组成及优势种
长江口北支4个年度底拖网调查共采集甲壳类34种,隶属于2目14科24属(表1)。虾类16种,蟹类17种,虾蛄类1种。年度差异方面,2012年捕获的甲壳类物种数最多,2011年捕获的甲壳类物种数最少,分别为23种和14种。渔获类别差异方面,虾类物种数以2011、2012和2013年最多,均为10种,以2014年最少,仅为6种;蟹类物种数以2012年最多,以2011年最少,分别为12种和3种;而虾蛄类在各年调查中均被捕获,且均为1种。
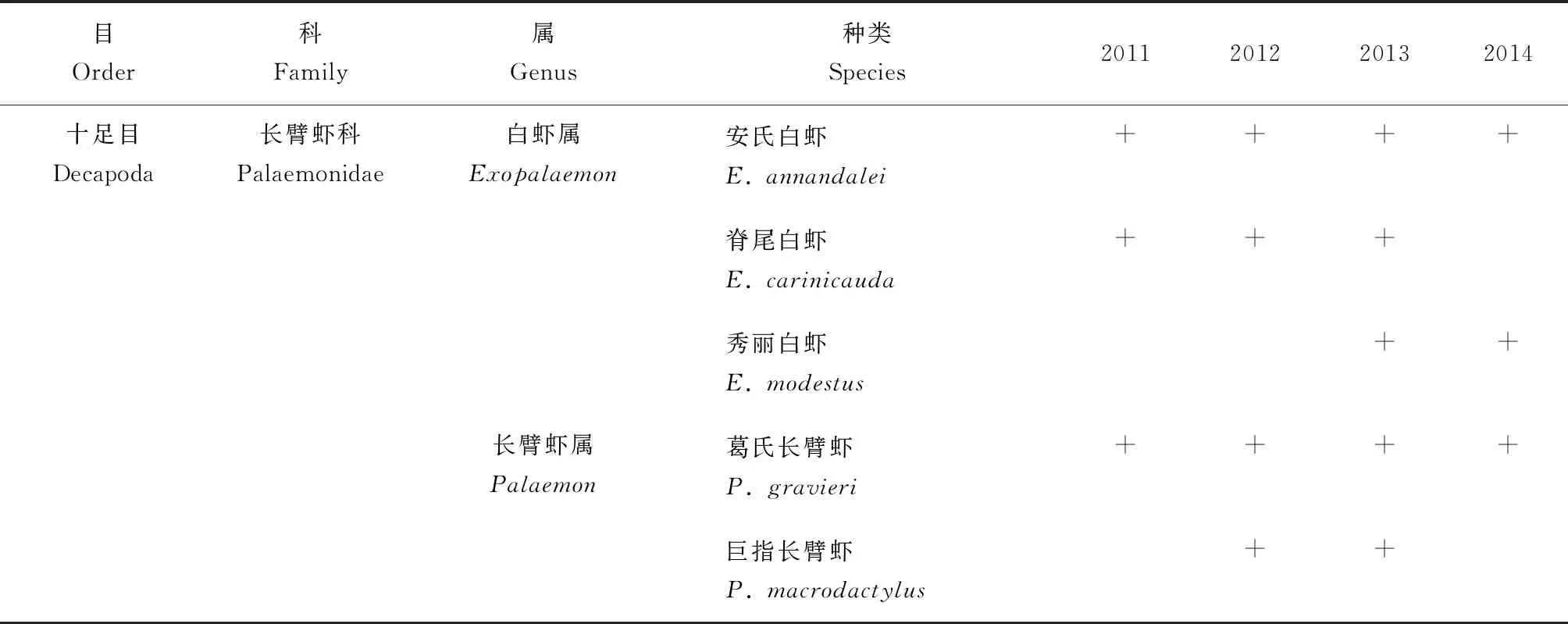
表1 2011─2014年长江口北支甲壳类种类组成

(续表1)

(续表1)
2011─2014年长江口北支各年度甲壳类优势种(Y≥0.02)变化情况见表2,调查期间共捕获优势种15种,其中虾类优势种8种,蟹类7种。葛氏长臂虾是长江口北支第一优势种,尤其是在2014年,优势度高达0.18。
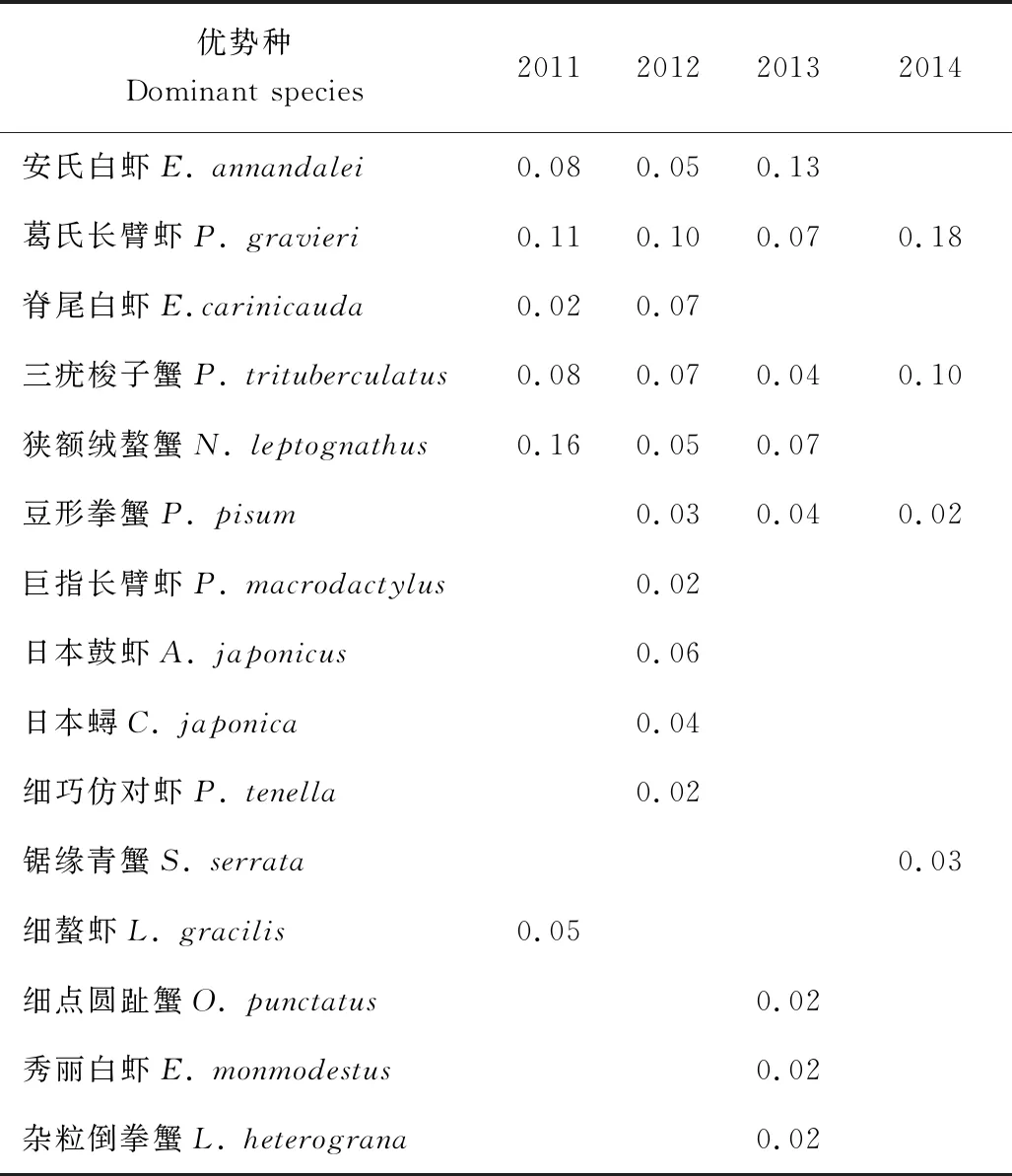
表2 2011─2014年长江口北支甲壳类优势种和优势度(Y≥0.02)
不同年度的优势种组成存在差异。葛氏长臂虾和三疣梭子蟹在4个年份均为优势种,年均优势度分别为0.12和0.07;安氏白虾、狭额绒螯蟹和豆形拳蟹在3个年份均为优势种,年均优势度分别为0.09、0.10和0.03;脊尾白虾在2个年份均为优势种,年均优势度为0.05。对比发现,单一年度优势种主要出现在2012年和2013年,在2011和2014年仅出现一种。这也表明在长江口北支4个年度中,以2012年优势种组成的种类最丰富,其群落结构相对较稳定。
2.2 资源密度组成
2011─2014年长江口北支甲壳类的平均生物量为11.43 kg/h(图2a)。其中以2012年甲壳类平均生物量最高,为19.53 kg/h;2014年甲壳类平均生物量最低,为3.47 kg/h。渔获组成差异方面,虾类平均生物量为1.405 kg/h,其中以2011年(1.63 kg/h)最高,2014年(1.03 kg/h)最低;蟹类的平均生物量为8.95 kg/h,其中以2012年(16.67 kg/h)最高,2014年(1.36 kg/h)最低;虾蛄类的平均生物量为1.075 kg/h,其中以2012年(1.35 kg/h)最高,2011年(0.75 kg/h)最低。
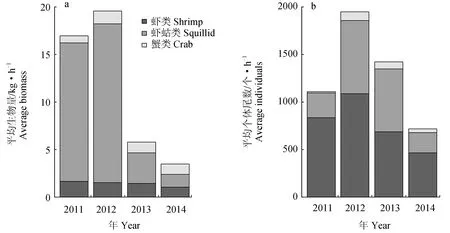
图2 2011─2014年长江口北支甲壳类资源密度指数的年际变化
2011─2014年,长江口北支甲壳类的平均个体尾数为1297.69个/h(图2b)。其中以2012年甲壳类平均个体尾数最高,为1944.77个/h;2014年甲壳类平均个体尾数最低,为717.89个/h。渔获组成差异方面,虾类的平均个体尾数为768.63个/h,其中以2012年(1087.56个/h)最高,2014年(463.65个/h)最低;蟹类的平均个体尾数为474.93个/h,其中以2012年(767.56个/h)最高,2014年(215.68个/h)最低;虾蛄类的平均生物量为54.13个/h,其中以2012(89.65个/h)年最高,2011年(13.78个/h)最低。
2.3 物种多样性
2011─2014年长江口北支各年度甲壳类物种多样性指数的变化规律较为一致(图3)。Shannon-Weiner多样性指数和Margalef丰富度指数的年度分布变化表现为先升后降的过程,且两者均值较为相近。其中,Shannon-Weiner多样性指数由2011年的0.83升至2012年的0.88,此时达到最高值,之后降至2014年的0.79,整体变化为0.79~0.88;Margalef丰富度指数由2011年的0.78升至2012年的0.87,此时达到最高值,之后降至2014年的0.81,整体变化为0.78~0.87。此外,Pielou均匀度指数的年度分布变化表现为先升后降再升的过程,其值由2011年的0.62升至0.66,此时达到最高值,之后降至2013年的0.64,最后再升至2014年的0.65,整体变化为0.62~0.66。经显著性分析显示,群落结构中物种多样性的年度差异较为显著(P<0.05)。

图3 长江口北支各年度甲壳类的物种多样性指数
2.4 群落结构分析
2.4.1 相似性
聚类分析结果(图4a)显示,在群落的相似性系数为21.4%时,长江口北支甲壳类组成被划分为2个组群,2011年、2013年和2014年共同称为第1组群,2012年单独称为第2组群。其中以2012年的群落结构相似性指数为最高值,其余年份的相似性指数均较低,表明其聚类结果与年度变化有关。根据非度量多维标度分析结果(图4b)中的胁迫系数值(Stress=0),其群落结构组成也被划分为2个组群,与聚类分析结果一致。这也说明了等级聚类图能够代表长江口北支甲壳类群落结构的排序。另外,相似性检验分析结果显示,各个组群间的群落结构组成均表现为显著性差异(P<0.05)。
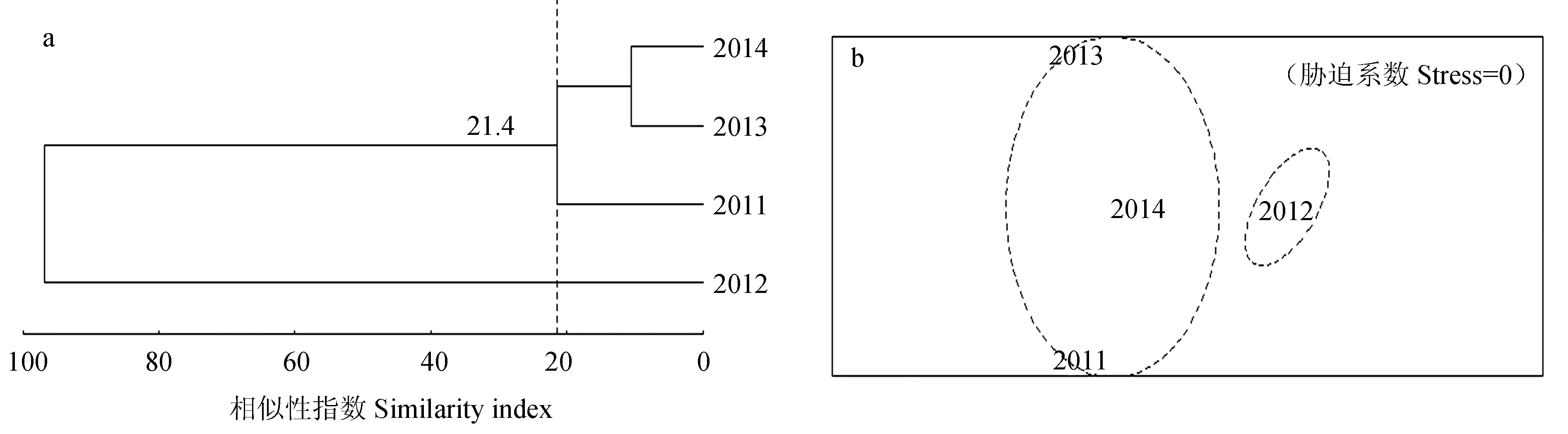
图4 长江口北支甲壳类群落的等级聚类及非度量多维标度排序分析
2.4.2 群落结构的稳定性
群落结构的更新指数和迁移指数,表征了其群落结构稳定性降低的节律(图5)。通过比较,2011─2014年长江口北支甲壳类群落的更新指数值表现为2014年>2011年>2013年>2012年,表明2012年和2013年群落的稳定性高于2011和2014年。
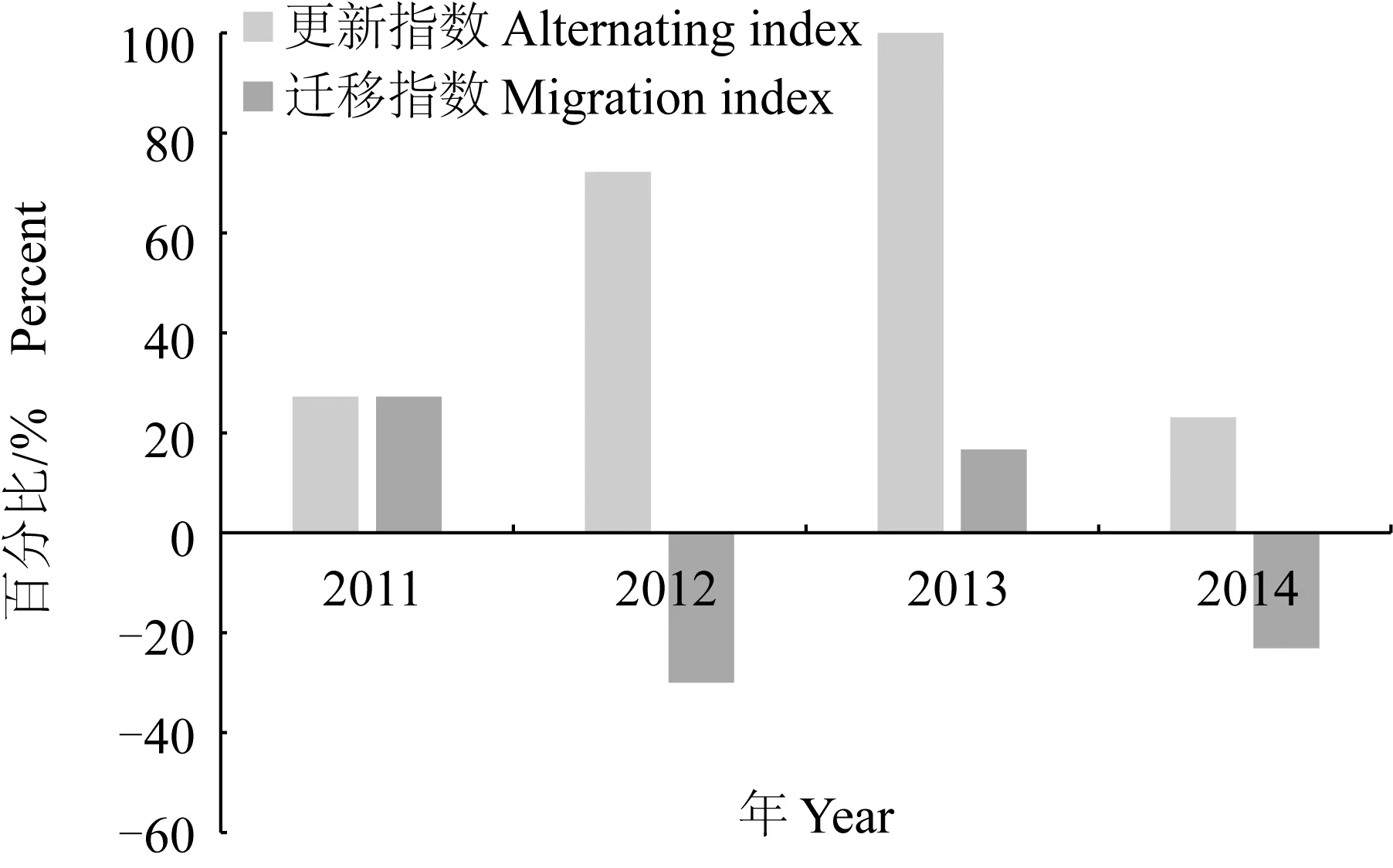
图5 2011─2014年长江口北支甲壳类群落的更新指数和迁移指数
不同年度甲壳类群落的迁移指数值存在一定差异,2012年和2013年迁入物种数大于迁出物种数,2011和2014年迁出物种数大于迁入物种数。由此发现,各年度其群落结构偏离动态平衡的程度不同。更新指数和迁移指数分析表明,2011年和2014年长江口北支甲壳类群落的稳定性最差,2012年和2013年其群落的稳定性最好。
通过丰度—生物量曲线的变化特征来衡量甲壳类群落结构的稳定性,一定程度上能反映出长江口北支生境遭受干扰或破坏的程度。2011和2014年的丰度曲线完全落在生物量曲线的上方(图6),表明这两个年度甲壳类的群落结构遭受了较为严重的扰动或破坏,群落结构稳定性较低。
相比较而言,2012年的生物量曲线完全在丰度曲线上方,表明甲壳类群落结构受到的扰动较小,群落结构比较稳定,而2013年的丰度/生物量曲线则存在重叠现象,丰度曲线基本出现生物量曲线的上方,表明2013年甲壳类的群落结构遭受了中等程度的扰动或破坏。
3 讨 论
3.1 甲壳类群落结构的种类组成及其优势种变化
长江口地处咸淡水交汇区域,受沿岸流、径流等影响较大,水文环境比较复杂,甲壳类物种一般多为广温广盐性[2,21]。本研究共捕获甲壳类物种34种,其中,葛氏长臂虾、三疣梭子蟹、脊尾白虾、安氏白虾和狭额绒螯蟹仍为全年常见优势种,其余多数物种只出现在单个年度。研究显示,通常纬度差异是影响物种分布的重要因素,一般高纬度海域的物种多样性低于低纬度海域[22]。对比不同海域,长江口北支捕获的甲壳类低于粤东柘林湾海域(43种)[23],高于唐山湾海域(17种)[24],但与渤海海域(33种)[25]捕获种类较为一致。此外,渤海海域甲壳类群落结构的年际变化比较明显,2012─2014年物种数目表现为先降后升的趋势,这与本研究结果不同。本研究中长江口北支4个年度甲壳类物种数目依次为14种、23种、21种和16种,物种数目表现为先升后降的趋势。这可能与不同海域调查面积、站点和海洋环境等有关[13-14]。
优势种代表群落结构中生物量或数量占据最高权重的生物种,优势种的变化一定程度上预示了该生物群落的演替[26]。长江口潮下带、滩涂等邻近海域研究发现[27-29],不同年度捕获的优势种类数目具有明显变化。本研究中,长江口北支共捕获优势种15种,优势种数目表现为2012年>2013年>2011年>2014年。此外,本次调查中,仅葛氏长臂虾和三疣梭子蟹在4个年度调查中均为优势种,表明其群落结构的优势种具有明显的年际变化,且与吴强等[25]研究结果一致。原因可能为长江口北支上游冲淡水作用导致盐度变化剧烈,适宜广温、广盐种类栖息[30],一定程度上也反映了长江口北支甲壳类群落结构的动态变化。
3.2 甲壳类群落结构资源密度组成的变化
已有研究表明,不同海域物种资源密度组成存在明显差异[31]。长江口及其邻近海域资源密度组成变化较大[14,28],这也与本研究结果观点相同。经本次调查发现,在长江口北支海域,甲壳类群落结构的资源密度年际变化整体表现为先升后降的趋势。如蟹类平均生物量最大,虾类次之,虾蛄类的平均生物量最小,分别为8.95、1.405 kg/h和1.075 kg/h,不同物种之间的平均生物量变化较大。经研究发现,除调查海域不同外,近几年长江口开展的增殖放流项目可能引起物种的资源密度组成发生变化[10,32]。本研究发现,2011─2014年长江口甲壳类物种组成匮乏,物种多样性较低,4个年度的平均甲壳类物种数目约为19种。陈强等[28]调查结果显示,长江口潮下带2013─2014年多样性指数整体较高,两个年度的平均甲壳类物种数为44种,远高于本研究结果。究其原因,这可能与研究区域有关[33],潮下带[28]、滩涂[34]和口外海域[12-13]等受上游来水的影响较小,而本研究区域属于口内海域,受到上游冲淡水影响较大,使得甲壳类物种数远低于陈强等[28]的研究结果。
然而,种类数及其均匀度也能够反映群落中的物种多样性[27,35],当群落结构和栖息环境受到不同程度的扰动时,如过度捕捞[27]、水域环境污染加剧[28]等,导致群落结构中的物种组成相对匮乏,表现为物种数目和生物量均呈现衰减趋势。
3.3 长江口北支生境遭受扰动或破坏的影响
对长江口北支甲壳类群落结构的研究,能够进一步认识其河口生态系统食物链中层营养级的重要性[25]。本研究中丰度—生物量曲线结果显示,长江口北支甲壳类的生物群落均受到不同程度的干扰,尤其是2011年和2014年受到的影响最大,其群落结构稳定性较差,物种多样性较低,而2012年遭受的扰动相对较小,群落结构相对稳定,2013年受到中等程度的扰动,这可能与“十二五”规划期间深水航道疏浚工程的验收及深水航道的回淤、减淤[36],以及北支盐水倒灌南支[37]有关,进而导致北支生境的不稳定性,使得群落多样性产生较大变动。
此外,物种栖息密度一定程度上也代表着群落结构的物种多样性,而物种栖息环境和受干扰程度等对长江口北支甲壳类的物种分布和栖息密度具有较大影响[27],这也是本次调查中影响生物多样性指数较低的重要原因。
4 结 论
通过对长江口北支水域的底拖网调查数据分析,本次调查共捕获甲壳类物种34种,葛氏长臂虾和三疣梭子蟹在4个年度中均为优势种,大多数物种仍为单年度优势种,且该水域优势种组成具有明显的年际变化;各年度甲壳类物种的资源密度组成不同,以2012年甲壳类的平均个体尾数和平均生物量值最高;经群落结构分析表明,不同组群间的群落结构组成及稳定性差异显著,其中,2012年生物量曲线完全在丰度曲线上方,表明其群落结构受扰动性较小,群落结构相对较稳定。综上,本研究结果对长江口北支水域生态环境修复及物种多样性保护等工作提供参考依据。
猜你喜欢
中国交通信息化(2022年7期)2022-10-27
当代水产(2022年8期)2022-09-20
法人(2022年3期)2022-03-30
昆明医科大学学报(2022年2期)2022-03-29
食品安全导刊(2021年20期)2021-08-30
福建林业科技(2020年4期)2020-12-31
今日农业(2020年19期)2020-12-14
安徽农学通报(2020年7期)2020-05-26
华东师范大学学报(自然科学版)(2019年2期)2019-06-11
水运管理(2017年10期)2017-11-20
