
植物非经典生长素信号转导通路解析
2020-08-04 03:02马军徐通达
生物技术通报 2020年7期
马军 徐通达
(福建农林大学海峡联合研究院 园艺植物生物学与代谢组学研究中心,福州 350007)
生长素英文名为“auxin”,取自希腊语“auxein”,意思是“生长”。生长素是最早被人们发现的一类植物激素,早在1880年,达尔文(Darwin)就发现植物的向光性生长依赖于其顶端产生的某种化学物质,该物质移动到下部组织并促使植物向光弯曲生长[1]。1926年,Went等[2]通过琼脂块扩散的方式从植物组织中分离到具有促进生长特性的化学物质。直到1946年,Haagen-Smit等[3]分离纯化并解析其化学结构为吲哚乙酸,是植物中生长素存在的主要形式。随后的大量研究发现生长素在植物生长发育的各个过程起着至关重要的调控作用[4-5]。植物主要通过协调生长素的合成代谢[6-7]、极性运输[8]以及信号转导[9]来实现其复杂多样的功能。
信号转导是将上游信号转换为下游复杂反应的重要过程。在细胞核内,泛素酶复合体关键蛋白TIR1/AFB蛋白被证明是生长素受体。生长素特异性地促进SCFTIR1/AFB-Aux/IAA蛋白复合体的形成从而导致Aux/IAA转录抑制因子的泛素化降解[10-13];经典Aux/IAA蛋白直接通过蛋白结合来抑制生长素响应转录因子ARF(Auxin response factor)最终调控下游基因转录[5,12]。在不同发育过程中,该信号通路通过不同组合的Aux/IAA-ARF转录调控元件,实现生长素调控的差异性和复杂性[5,14-16]。TIR1/AFB介导的生长素转录调控机制被称为经典生长素信号通路[14,17]。然而,植物生长素的复杂功能不能完全通过TIR1/AFB介导的经典信号通路来解释,例如生长素的快速反应、细胞间的生长素响应以及亚细胞水平上的调控等[14,17-19]。近期研究逐步阐明生长素在不同调控水平上存在非经典信号传递机制,通过与经典信号转导通路的相互配合,协同控制植物复杂的发育过程[14,17-19]。
1 TIR1/AFB介导的非经典生长素信号转导通路
1.1 生长素调控细胞伸展的分子机制
生长素促进下胚轴细胞的细胞伸展(Cell expansion)是典型的生长素功能之一。前期研究发现生长素主要通过细胞质膜质子泵ATP酶(PM H+-ATPase)的激活和质外体(Apoplast)的酸化来促进植物细胞生长,也就是 “酸性生长假说”[20]。细胞生长的动力来自于细胞内液泡的膨压,同时需要细胞壁结构软化来为细胞生长提供空间。酸性生长调控机制的核心元件是定位在细胞膜上的质子泵蛋白H+-ATPase。生长素通过激活H+-ATPase降低细胞壁的PH值,通过调控细胞壁相关蛋白包括Expansin蛋白、XTH(Xyloglucan endotransglycosylase)蛋白以及PME(Pectin methylesterases)蛋白,增加细胞壁的扩展性能,从而促进细胞扩展[20]。生长素酸性生长假说的核心问题是生长素如何激活质膜定位蛋白H+-ATPase。研究发现拟南芥下胚轴细胞的酸性生长依赖于TIR1/AFB-Aux/IAA-ARF信号通路[21]。生长素通过TIR1/AFB信号通路局部激活H+-ATPase从而介导细胞壁的快速酸化和下胚轴的伸长,该结果为生长素酸性生长假说提供了直接的实验证据[21]。除此之外,研究发现在拟南芥中,AHA2(Autoinhibited H+ATPase2)蛋白的碳末端存在关键的调控位点,其Thr947位点的磷酸化决定了其质子泵的活性[22]。生长素能在10 min内快速磷酸化AHA2蛋白的Thr947位点从而激活该蛋白,进而调控细胞壁的酸碱性从而调控细胞生长[22]。为了研究生长素快速磷酸化激活AHA2蛋白是否通过核内受体TIR1/AFB蛋白,研究人员根据TIR1受体的蛋白结构,通过工程学方法将TIR1蛋白改造为ccvTIR1(concave TIR1)蛋白,改造后的ccvTIR1只能特异性识别改造后的生长素cvxIAA(convex IAA)而不能识别未改造的IAA,从而规避生长素的其他复杂响应来特异性研究TIR1受体介导的信号通路[23]。有意思的是,改造后的生长素cvxIAA在ccvTIR1转基因植物中依然能够快速促进AHA2蛋白的磷酸化,这说明该细胞膜上的生长素快速反应依然部分依赖于细胞核TIR1/AFB受体[23]。然而到目前为止,TIR1/AFB受体直接参与该快速反应的分子机制仍不清楚。
生长素通过磷酸化修饰调控AHA的蛋白活性,意味着一定有激酶和磷酸酶参与到该调控过程。研究发现,磷酸酶家族蛋白D分支的PP2C.D蛋白能够直接和AHA2蛋白互作并调控其Thr947位点的去磷酸化,从而影响植物细胞生长。生长素通过经典TIR1/AFB信号通路诱导SAUR(Small Auxin Up RNA)基因表达,SAUR蛋白的积累能够竞争性结合PP2C.D磷酸酶蛋白来抑制该磷酸酶对AHA2的去磷酸化从而激活AHA2[24]。该机制部分揭示了生长素如何调控H+-ATPase的活性,然而到目前为止直接参与AHA2磷酸化修饰的蛋白激酶仍有待于进一步的寻找和探索(图1)。
1.2 生长素快速抑制根生长的信号通路
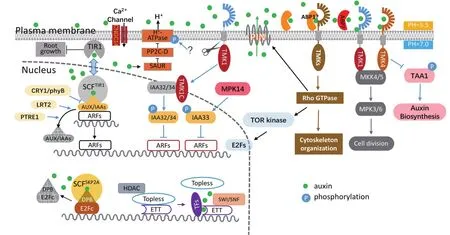
图1 拟南芥经典和非经典生长素信号通路示意图
生长素对植物根器官生长的快速抑制是另一个典型的生长素响应过程,其分子机制仍不清楚。最近的研究利用微流控技术、垂直活体显微成像技术和数学模拟等方法[25-27],探索了生长素快速抑制植物根系伸长的分子机制[28]。当利用微流控技术对拟南芥根系施加生长素时,生长素对根的抑制反应非常迅速,发生在30 s以内;当去除生长素时,根系生长快速恢复,发生在2 min以内,几乎没有滞后期。如此快速的生长素反应似乎无法通过细胞核内生长素TIR1受体介导的经典转录调控机制来解释,然而通过对tir/afb突变体进行分析以及对cvxIAA-ccvTIR1配对的生理效果进行检测[23]发现TIR1/AFB受体对于生长素快速调控根系生长是必需的,证明生长素TIR1/AFB受体可能存在非转录调控的分子机制,这还有待于进一步发掘。
生长素快速抑制植物根系生长的另一个机制是通过细胞膜上钙离子的瞬间跨膜流动,导致细胞内外的钙离子浓度和PH的变化来实现的[29]。研究发现生长素处理和重力刺激通过环核苷酸门控通道蛋白 CNGC14(Cyclic Nucleotide-gated Channel) 引 起钙离子的快速流入和膜外碱化,细胞壁碱化导致细胞伸长被抑制,因此在cngc14突变体中生长素引起的生长抑制和向地性弯曲生长都显著延迟[30]。生长素10 s内引起的钙离子流动显然也难以用生长素经典转录调控机制来解释。最近,Dindas等[31]发现根毛中生长素内运蛋白AUX1(Auxin resistant 1)对于根毛生长素摄取和膜去极性化是必须的;AUX1介导的生长素运输以及生长素触发的钙信号能够被TIR1/AFB抑制剂auxinole阻断;此外,在tir1afb2afb3和cngc14突变体中,AUX1介导的生长素运输以及生长素触发的钙信号变化也显著降低。这说明AUX1转运蛋白、TIR1/AFB受体与CNGC14钙离子通道都参与调控根部生长素的快速响应过程。生长素能够在数秒内诱导根毛质膜的钙离子内流[31],而且这种效应依赖于生长素内运蛋白AUX1以及TIR1/AFB受体,再一次证明TIR1/AFB受体可能存在转录调控之外的分子机制[32](图1)。
2 不依赖于TIR1/AFB的非经典生长素信号转导通路
2.1 非经典ARF介导的生长素信号转导通路
经典的生长素响应转录因子ARF家族蛋白通过其PB1结构域与Aux/IAA蛋白结合,从而被TIR1/AFB受体调控。然而ARF家族蛋白中存在非经典ARF(图2-A),如拟南芥ARF3蛋白(又被称为ETTIN蛋白),因缺失PB1结构域而不能与经典TIR1/AFB信号通路的元件相互作用,因此ETTIN的功能并不依赖于TIR1/AFB受体[17]。研究发现ETTIN蛋白的C末端是其功能调控的关键区域,具有丝氨酸富集的特点,因此可能被上游激酶调控[33]。同时ETTIN蛋白能和其他的转录调控因子包括 INDEHISCENT(IND)、REPLUMLESS(RPL)、BREVIPEDICELLUS(BP)蛋白互作,而生长素能够抑制这些蛋白复合体的形成,从而调控基因转录[34-36]。有意思的是,最新的研究发现生长素能够直接与ETTIN蛋白特异性结合,抑制ETTIN蛋白和TOPLESS蛋白与HDA9组蛋白乙酰转移酶的互作,促进组蛋白乙酰化,调控基因转录的重编程,从而参与植物关键发育过程[37-38]。这也意味着生长素能够直接通过非经典ARF调控基因转录。这其中还有许多疑问有待进一步解析,例如ETTIN蛋白是否是新的生长素受体;ETTIN蛋白介导的生长素非经典信号通路如何与经典的TIR1/AFB-AUX/IAA-ARF信号通路相互协调控制基因转录等(图1)。

图2 拟南芥ARF蛋白和IAA蛋白的系统进化树分析
2.2 非经典Aux/IAA介导的生长素信号转导通路
经典的转录抑制因子Aux/IAA蛋白存在4个结构区域,其中蛋白结构域II(DII)是Aux/IAA与TIR1/AFB 受体结合的关键区域[15,39]。生长素通过促进TIR1/AFB和Aux/IAA的蛋白互作,将经典Aux/IAA蛋白多聚泛素化,从而通过蛋白酶降解。除此之外,经典IAA也存在别的调控模式,其中包括光受体PhyB/CRY1通过竞争性抑制TIR1/AFB受体与Aux/IAA的互作来调控生长素响应;LRT2(Lateral rootless 2)通过调控Aux/IAA蛋白的顺反异构来决定其与TIR1/AFB受体的互作;PTRE1(Proteasome Regulator 1)通过抑制蛋白酶体的活性来调控Aux/IAA的降解[40-42],这些重要发现都充分证明了Aux/IAA蛋白调控的复杂性。Aux/IAA蛋白家族中也存在一类非经典Aux/IAA蛋白,因其蛋白缺失结构域II而可能不被TIR1/AFB受体识别,这其中包括拟南芥 IAA20、IAA30、IAA31、IAA32、IAA33 和 IAA34 蛋白[15,39](图 2-B)。由于非经典IAA蛋白无法被TIR1/AFB调控,因此其调控和作用机制有别于经典IAA蛋白。最近一系列研究发现非经典IAA蛋白的作用机制与经典IAA蛋白完全不同,生长素非但没有通过TIR1/AFB降解非经典IAA蛋白,反而通过上游蛋白激酶磷酸化修饰稳定非经典IAA蛋白[43-44]。在子叶顶端弯钩发育过程中,弯钩内侧生长素通过TMK激酶调控非经典IAA32和IAA34蛋白的稳定性,然后通过ARF转录因子调控基因表达来维持子叶弯钩内外侧的差异性生长[43]。在根尖分生组织,生长素通过MPK14调控非经典IAA33蛋白的稳定性,从而和经典IAA5蛋白竞争性结合下游ARF10和ARF16,调控基因转录来维持根尖分生组织的稳态[44]。上述研究阐明了非经典IAA蛋白不再被TIR1/AFB介导的泛素化蛋白降解途径调控而是被上游相关激酶途径调控;与此同时生长素不再降解非经典IAA蛋白反而促进其蛋白的稳定性;非经典IAA蛋白仍然通过ARF转录因子调控基因转录。对IAA蛋白的进化树研究发现,非经典IAA在进化上也很保守,这意味着在植物自然选择和进化过程中,非经典IAA的功能也是必须的。非经典IAA蛋白介导的生长素信号通路仍有许多未知的问题有待于解决,如非经典IAA蛋白既然不通过TIR1/AFB泛素化途径降解,那其降解机制是什么;其他非经典IAA蛋白的功能和调控机制也有待于进一步解析(图1)。
2.3 小G蛋白ROP GTPase介导的非经典生长素信号通路
植物ROP(Rho-related GTPase of Plant)蛋白信号途径是生长素非经典信号通路的另外一个重要分支。ROP蛋白被认为植物信号转导的分子开关之一,存在GTP结合的活性形式和GDP结合的非活性形式。当ROP蛋白被激活之后,可以开启下游复杂的信号调控网络,调控包括细胞骨架、极性运输、基因转录等过程[45]。研究发现,生长素能够快速激活ROP蛋白。在叶表皮细胞中,生长素激活ROP2蛋白调控微丝蛋白动态,激活ROP6调控微管蛋白动态,从而调控叶表皮细胞的极性建成和形态发生。在根毛发育过程中,生长素通过小G蛋白在生毛细胞中的极性定位决定了根毛产生的位置[46-47]。同时,对ROP活性的调控也影响根毛产生的位置和生长,如过表达ROP2会产生更多且更长的根毛[47]。最新研究发现,生长素能够通过小G蛋白ROP2激活TOR(Target of Rapamycin)激酶,然后通过E2F转录因子,调控植物分生组织的细胞分裂活性[48]。然而,生长素激活小G蛋白非常迅速,在30 s就能激活小G蛋白ROP2和ROP6[49-51],这种快速激活意味着生长素激活小G蛋白的机制不可能依赖于TIR1/AFB介导的转录调控,而存在新的调控机制。
研究发现生长素结合蛋白ABP1(Auxin binding protein 1)以及类受体激酶家族蛋白TMK(Transmembrane Kinases)介导生长素对小G蛋白的快速激活[49-50]。ABP1蛋白因早期T-DNA突变体遗传材料的问题,其功能有待于进一步解析[52-54]。近期研究发现,生长素能够促进小G蛋白ROP6在细胞膜上与膜磷脂结合形成微聚体来激活ROP6信号通路[55],而这个过程也依赖于 TMK1 和 TMK4[56]。这些研究说明,生长素调控小G蛋白是发生在细胞膜上的分子水平反应,其原理的进一步解析有助于理解生长素快速反应的产生机制(图1)。
2.4 TMK家族蛋白介导的生长素信号转导机制
类受体激酶通常是传递信号的关键元件,在植物激素信号转导中具有极其重要的作用,如类受体激酶BRI1(Brassinosteroid insensitive 1)作为油菜素内酯受体介导其信号转导过程[57]。最近研究发现类受体激酶家族TMK蛋白(Transmembrane Kinase)在生长素信号转导中发挥着至关重要的作用,从非转录水平调控植物细胞的生长素响应[43,58-59]。除了生长素激活小G蛋白信号通路依赖于TMK家族激酶,在侧根发育过程中,局部有序的生长素分布通过TMK1和TMK4调控器官发生过程中细胞分裂模式的有序性,从而精准调控侧根的有序发生[58]。研究发现生长素通过TMK1/4磷酸化激活丝裂原活化蛋白激酶(Mitogen-activated protein kinase)MKK4/5-MPK3/6信号途径,来调控细胞分裂模式,也是生长素信号和MPK信号途径直接关联的重要证据[58]。最新研究发现,生长素能够通过TMK4蛋白磷酸化修饰生长素合成关键酶TAA1(TRYPTOPHAN AMINOTRANSFERASE OF ARABIDOPSIS),负调控生长素的生物合成[59]。这也是首次报道生长素合成的翻译后调控机制,说明植物体内生长素浓度存在快速的自我调控机制,以满足植物生长发育的需求[59]。
有意思的是,TMK家族蛋白不仅参与生长素的非转录水平调控,也参与转录水平的调控。研究发现高浓度生长素能够激活一条全新的抑制植物局部生长的信号通路。在子叶顶端弯钩内侧聚集的高浓度生长素能够促进TMK1蛋白的剪切,通过剪切下来的TMK1蛋白的C末端肽段将生长素信号传递到细胞质和细胞核调控下游反应。TMK1蛋白C端特异性识别并磷酸化稳定非经典IAA蛋白IAA32/34,IAA32/34蛋白的积累通过结合ARF转录因子来抑制基因转录,最终导致在子叶顶端弯钩内侧细胞伸长被抑制来维持内外侧差异性生长[43]。说明生长素TIR1/AFB和TMK1介导的生长素信号途径精准分工,分别选择经典和非经典的IAA蛋白进行差异化调控下游反应。
拟南芥中TMK家族蛋白有4个成员,不同的TMK蛋白在植物生长发育的不同阶段表达,从而行使不同的功能。拟南芥tmk1;tmk2;tmk3;tmk4四突变体表现极其严重的发育缺陷,最终导致不育的表型,说明TMK家族蛋白功能的复杂性和多样性[60]。研究已经表明,TMK激酶介导的生长素信号转导通路在不同植物组织、不同亚细胞位置(从细胞膜到细胞核)、不同水平上(从非转录水平到转录水平)全方位调控植物生长发育。然而,TMK家族介导的生长素信号转导机制仍有许多未解决的问题,例如生长素信号是如何被TMK类受体激酶感知?TMK的作用是否需要其结合蛋白ABP1?TMK家族蛋白剪切介导的信号传递机制是否普遍存在及调控机制是什么?这些问题都有待于进一步解析。
3 展望:非经典生长素信号途径的研究瓶颈和前景
本综述初步概括了生长素非经典信号传递通路的研究进展。生长素调控植物生长发育极其复杂和多样。这些研究进展一方面拓展了对生长素调控网络的深入认识,另一方面也提出了新的问题和挑战。
3.1 非经典信号通路中生长素的感受机制
随着非经典信号通路的逐步解析,生长素是如何被感知的依旧是核心问题之一。其中包括:(1)生长素TIR1/AFB受体是否存在其他不依赖于Aux/IAA蛋白的作用机制。(2)TMK家族激酶是否能够像BRI1一样直接感知生长素,是否需要类似ABP1的辅助蛋白。(3)生长素结合蛋白ABP1是否参与非经典生长素信号通路;abp1单突变体表型不明显,这和ABP1生化和细胞水平功能相矛盾,如何去解析;ABP1作为没有功能域的结构蛋白,是否存在相似结构蛋白的功能冗余或功能补偿效应。(4)是否存在别的生长素结合蛋白,如何去寻找和鉴定。
3.2 生长素经典与非经典信号通路的协同调控
生长素作为小分子化合物在植物细胞内聚集后,同时启动经典和非经典信号通路来调节植物的生长发育,必然存在精准的协同机制。例如,多信号通路之间在时间和空间上是如何协同的,如何解析正反馈调控与负反馈调控的复杂效应;多信号通路在不同组织器官对不同浓度生长素如何协同产生不同的发育效应的;多信号通路如何协同参与植物对复杂环境的响应的。这些问题的解答,将有助于全面解析生长素信号调控网络,为精准调控生长素响应奠定理论基础。
猜你喜欢
云南医药(2021年3期)2021-07-21
生物学通报(2021年4期)2021-03-16
科学大众(2020年12期)2020-08-13
学苑创造·A版(2020年12期)2020-01-07
中国外汇(2019年15期)2019-10-14
数码世界(2018年7期)2018-12-24
作文教学研究(2016年1期)2016-07-05
医学研究杂志(2015年8期)2015-06-22
医学研究杂志(2015年4期)2015-06-10
中国医药导报(2015年24期)2015-02-28
